
A revision of the egg-eating snakes of the genus Dasypeltis Wagler (Squamata: Colubridae: Colubrinae) in north-eastern Africa and south-western Arabia, with descriptions of three new species
Michael F. Bates1 & Donald G. Broadley2†
1Department of Herpetology, National Museum, P.O. Box 266, Bloemfontein 9300, South Africa
2 Department of Herpetology, Natural History Museum of Zimbabwe, P.O. Box 240, Bulawayo, Zimbabwe
1 Corresponding author: E-mail: herp@nasmus.co.za
Bates & Broadley – Revision of Dasypeltis in NE Africa and SW Arabia
CONTENTS
Abstract
Introduction
Materials and Methods
Character analysis
Systematics
Dasypeltis medici (Bianconi)
Dasypeltis confusa Trape & Mané
Dasypeltis scabra (Linnaeus)
Dasypeltis bazi Saleh & Sarhan
Dasypeltis taylori Bates & Broadley sp. nov.
Dasypeltis atra Sternfeld
Dasypeltis fasciata A. Smith
Dasypeltis abyssina (Duméril, Bibron & Duméril)
Dasypeltis crucifera Bates sp. nov.
Dasypeltis arabica Broadley & Bates sp. nov.
Multivariate analyses
Summary of species
Discussion
Key to the genus Dasypeltis in north-eastern Africa and south-western Arabia
Acknowledgements
References
Appendix
Abstract
The genus Dasypeltis Wagler, 1830 in north-eastern Africa (from latitude 8ºS to the Mediterranean Sea, east of 29°E) and south-western Arabia (west of longitude 45°E) is reviewed. Ten species are recognised in this region on the basis of morphological characters, including colour pattern. Dasypeltis fasciata A. Smith, 1849 inhabits lowland forest in western and southern Uganda, while Dasypeltis medici (Bianconi, 1859) occurs in coastal forest/savannah mosaic in southern Somalia, Kenya and northern Tanzania. We show that Dasypeltis medici lamuensis Gans, 1957 is a junior synonym of D. medici. Dasypeltis atra Sternfeld, 1912—in a variety of colour phases/patterns—is widespread in the region, occurring in montane forest, moorland, forest/savannah mosaic, and savannah along the border between Tanzania and Kenya. Dasypeltis scabra (Linnaeus, 1758) is widely distributed in East African savannahs, except in parts of South Sudan, Uganda, western Kenya and Rwanda where Dasypeltis confusa Trape & Mané, 2006 infiltrates from the west, and in northern Somalia and Djibouti between 600 and 1370 m where it is replaced by Dasypeltis taylori Bates & Broadley sp. nov. Dasypeltis bazi Saleh & Sarhan, 2016 occurs as an isolated population in the Fayoum Depression west of Cairo, Egypt, with a possible isolate at Erkowit in the Red Sea Hills of north-eastern Sudan. We designate a lectotype for Dasypeltis abyssina (Duméril, Bibron & Duméril, 1854)—previously known only from the type description—and demonstrate that it is a distinct and valid species restricted to open savannah in the highlands (1800–2450 m) of north-western Ethiopia and central Eritrea; the paralectotype from Sudan is referable to D. scabra. Dasypeltis crucifera Bates sp. nov. is known only from two localities at lower elevations (600–1417 m) in northern Eritrea. Dasypeltis arabica Broadley & Bates sp. nov. inhabits the highlands of south-western Arabia (Saudi Arabia and Yemen) at elevations of 1600–2200 m. Multivariate analyses of scale characters and numbers of pattern cycles provides support for the above taxonomy. We provide a map detailing the ranges of the various species. The Horn of Africa, with at least six species (and another in Arabia), appears to be a centre of diversity for the genus Dasypeltis.
Key words: Dasypeltis, taxonomy, morphology, zoogeography, north-eastern Africa, south-western Arabia
† Donald G. Broadley passed away on 10 March 2016.
INTRODUCTION
Carl Gans (1959) noted that the egg-eating snakes of the genus Dasypeltis Wagler 1830 had attracted considerable attention because of their exclusive diet of bird eggs and the related adaptations of their skull and anterior vertebrae, but that identification of specimens was often difficult. This, he suggested, was because previous analyses had been limited to relatively small areas—and therefore geographical variation in colour pattern and scalation features was poorly understood. With this in mind he conducted a detailed taxonomic analysis on a large sample (over 1000 specimens) representing the entire range of the genus, i.e. sub-Saharan Africa, Egypt and south-western Arabia. His aim was firstly to distinguish valid species, and secondly to describe geographical and individual variation within each of these. Because of a scarcity of material from several large areas, his emphasis was on describing variation rather than attempting to validate existing subspecies or describe new taxa, and his taxonomic conclusions were therefore deliberately conservative. Gans (1959) found that colour pattern was a very useful character and used this, in combination with various other morphological features, to separate species and distinguish populations.
Gans (1959) identified nine basic dorsal colour patterns, some with sub-types. Most of these patterns, including a few of the sub-types, are now considered diagnostic of various species. For the widespread Dasypeltis scabra (Linnaeus, 1758) he documented a few distict variations in dorsal colour pattern. Some of these patterns were largely or wholly restricted to particular regions. For example, Gans’ (1959) ‘brown’ grouping included, for central and eastern Africa, three types: brown (‘2B’), black (‘2M’) and marked (‘2Bx’; typically with dark bands—often incomplete—across the back), these being referable to Dasypeltis atra Sternfeld, 1912 (see also Gans 1964; Broadley & Bates 2009). Specimens with the linked ‘5L’ colour pattern, consisting of dark, more-or-less diamond-shaped saddles confluent with dark lateral bars—such that they formed bands over the body—occur in western and central Africa, and these populations are now referable to Dasypeltis confusa Trapé & Mané, 2006 (see also Bates 2013). Another variation was ‘5I’ where black rectangular saddles and lateral bars form complete closure around the pale inter-saddle areas. However, most other populations of D. scabra throughout its extensive range had the variable non-linked ‘5N’ dorsal pattern consisting of dark markings in the form of large ‘saddles’ (varying in size and shape) along the middle of the back, alternating with lateral bars.
In his taxonomic revision of Dasypeltis, Gans (1959) recorded only three species from north-eastern Africa: the widespread D. scabra, two subspecies of Dasypeltis medici (Bianconi, 1859)—Dasypeltis medici medici (pattern ‘8’: usually dark rectangular saddles separated by narrow pale interspaces, with adjacent narrow dark lateral bars) in southern Kenya and Tanzania, and Dasypeltis medici lamuensis Gans, 1957 (pattern ‘3’: usually unmarked or faintly marked) along the coast from Somalia to the Kenya-Tanzania border, and Dasypeltis fasciata A. Smith, 1849 (pattern ‘4’: checker board pattern on back) which occurred marginally in western Uganda. However, in 1964 Gans recognised Dasypeltis atra Sternfeld, 1912 as a full species in the montane forests of the region. Broadley & Howell (1991), in a key to the Tanzanian species of Dasypeltis, separated ‘2Bx’ patterned specimens of D. atra (see Gans 1959, 1964) from D. scabra on the basis of their higher numbers of pattern cycles (85–107 versus 42–79). Largen & Rasmussen (1993) recognised only a single species of egg-eater, namely D. scabra, in Ethiopia and Eritrea. Hughes (1997) mapped the distribution of Dasypeltis in Uganda, based on material in the Natural History Museum (London) and the World Museum (Liverpool), but doubted whether the ‘2Bx’ colour phase of Gans (1959) should be assigned to D. atra.
In their revision of West African Dasypeltis, Trape & Mané (2006), in an aside, revived Dasypeltis abyssina (Duméril, Bibron & Duméril, 1854) of Ethiopia from the synonymy of D. scabra, after examining the syntypes in Paris. However, they provided only scant details as to why they considered this taxon valid. Broadley & Bates (2009) documented range extensions of the patterned (‘2Bx’) phase of D. atra into savannah areas of northern Tanzania, and the dorsally uniform black and brown phases into south-western Tanzania. Largen & Spawls (2010) recorded D. scabra from Ethiopia and Eritrea, and D. atra from Ethiopia, but also illustrated a D. abyssina (labelled as D. scabra, see below) from Mulu (actually at Keriyo hamlet, S. Spawls in litt. 5 April 2011) where it is sympatric with D. atra. An isolated population in the Fayoum Depression in Egypt was recently described as Dasypeltis bazi Saleh & Sarhan, 2016.
There are currently 13 recognised species of Dasypeltis, namely D. scabra, D. palmarum (Leach), D. inornata Smith, D. fasciata, D. abyssina, D. medici (D. m. medici and D. m. lamuensis), D. atra, D. confusa, D. gansi Trape & Mané, 2006, D. latericia Trape & Mané, 2006, D. sahelensis Trape & Mané, 2006, D. parascabra Trape, Mediannikov & Trape, 2012, and D. bazi (see Gans 1959, 1964; Trape & Mané 2006; Trape et al. 2012; Saleh & Sarhan 2016).
Since Gans’s (1959) revision, significant collections of specimens have become available for most parts of the study area, and this allowed for a more detailed evaluation of variation and patterns of geographical distribution. Several taxonomic problems for populations in north-eastern Africa and south-western Arabia require resolution. For example, there has been much uncertainty about the status and distribution of D. atra due to the wide variety of colour morphs and confusion with patterned D. scabra (see Hughes 1997; Broadley & Bates 2009). Examination of most of the available Dasypeltis material from this region also yielded new records of D. abyssina—a species which until now has not been properly diagnosed from its congeners—and recognition of easily identifiable and apparently closely related populations from the Keren-Agordat area, and across the Red Sea in the highlands of the south-western Arabian Peninsula, both described here as new species. In addition, an isolated population in Somaliland and Djibouti is described as a new species.
MATERIALS AND METHODS
Study area.—The study area is defined as Egypt, other parts of Africa north of latitude 8° S and east of longitude 29° E, and the Arabian Peninsula west of longitude 46° E. It therefore includes all eastern populations of D. atra, and the eastern-most populations of D. confusa and D. fasciata. To the north the area extends to the Mediterranean Sea and encompasses the isolated Egyptian population of D. bazi (see Gans 1959; Saleh & Sarhan 2016). The countries and regions covered by this revision are therefore: south-western Saudi Arabia, western Yemen, Egypt, the eastern parts of Sudan and South Sudan, Eritrea, Ethiopia, Djibouti, Somalia, Uganda, Kenya, Rwanda (which includes a small area west of 29° E), Burundi and northern Tanzania.
Material examined and characters used.—The starting point for this study was the relevant scalation, size and colour pattern data published in Appendix 1 of Gans (1959), together with the appropriate plates. Data in Gans (1964: 293) was also used. Most of the relevant material in the Natural History Museum in London was examined by us, together with several additional important specimens in other European, North American and African museums. The institutional abbreviations used below are from Sabaj (2016), except we use ‘BM’ for Natural History Museum in London, ‘MUZM’ for Makerere University Zoology Museum in Kampala, ‘AJL’ for the private collection of Angelo Lambiris in Durban (to be incorporated into the Durban Natural Science Museum), and ‘CG’ refers to the Carl Gans collection (see Gans 1964).
AJL Angelo Lambiris private collection, Durban
AMNH American Museum of Natural History, New York
AAU Addis Ababa University, Department of Biology
BM Natural History Museum, London
CAS California Academy of Sciences, San Francisco
CG Carl Gans collection (Gans 1964)
CM Carnegie Museum of Natural History, Pittsburgh
FMNH Field Museum of Natural History, Zoology Department, Chicago
IRSNB Intitut Royal des Sciences Naturelles de Belgique, Brussels
LACM Natural History Museum of Los Angeles County, Los Angeles
LIV World Museum, Liverpool
MCZ Museum of Comparative Zoology, Harvard University, Cambridge
MHNG Muséum d’Histoire naturelle, Genève [Geneva]
MNHN Muséum national d’Histoire naturelle, Paris
MRAC Musée Royal de l’Afrique Centrale, Tervuren
MSNG Museo Civico di Storia Naturale “Giacomo Doria”, Genova [Genoa]
MSNM Museo Civico di Storia Naturale di Milano [Milan]
MSNPV Museo di Storia Naturale dell’Università di Pavia [Pavia]
MUZM Makerere University Zoology Museum, Kampala
MZUF Museo di Storia Naturale di Firenze, Sezione di Zoologia “La Specola”, Firenzi [Florence]
MZUT Museo di Zoologia, Università di Torino [Turin]
NMBO National Museum, Bloemfontein
NMK National Museums of Kenya, Nairobi
NMW Naturhistorisches Museum, Wien [Vienna]
NMZB Natural History Museum of Zimbabwe, Bulawayo
PEM Port Elizabeth Museum, Bayworld
RMNH Naturalis Biodiversity Centre, Leiden
TMP Ditsong National Museum of Natural History, Pretoria
SMNS Staatliches Museum für Naturkunde, Stuttgart
TCWC Biodiversity Research and Teaching Collections, Department of Wildlife and Fisheries Sciences, Texas A&M University, College Station
UMMZ University of Michigan Museum of Zoology, Ann Arbor
USNM National Museum of Natural History, Smithsonian Institution, Department of Vertebrate Zoology, Washington D.C.
ZFMK Zoologisches Forschungsmuseum Alexander Koenig, Bonn
ZMB Museum für Naturkunde, Berlin
ZMUC Kobenhavns Universitet, Zoologisk Museum [Zoological Museum, University of Copenhagen], Copenhagen
Scalation and size data was also obtained from the curators of various museums (see Acknowledgements) and B. Hughes (for Ugandan material in the World Museum, Liverpool [LIV]). In several cases we recorded or confirmed the dorsal, and in some cases also ventral, colour patterns of specimens by examining colour images of specimens from various museums (e.g. World Museum, Liverpool; Museo Civico di Storia Naturale “Giacomo Doria”, Genova [Genoa]; Museo Regionale di Scienzi Naturali, Turin; National Museums of Kenya, Nairobi). Data is available for a total of 640 specimens (Appendix 1).
Material listed under ‘Localities’ comprises specimens examined by the authors, or for which data and/or photographs were provided and allowed positive determinations. In this section countries are listed from north to south, while specific localities are listed alphabetically. The mapping notation (e.g. SE 0135B3) refers to quarter-degree units (e.g. Minter et al. 2004: 48; but for the last part of the code we use numbers 1 to 4 instead of letters A to D), although on the map (Fig. 3) we plot these as half-degree units.
Most of the characters used by Gans (1959) were employed and evaluated in the same way in the present study, with modifications (or explanations) as indicated below. Specimens were examined under a binocular dissecting microscope or with the aid of magnifying lenses for some of the larger specimens. Ventrals were counted from the first scale following the posterior pair of chin shields, up to but excluding the anal plate (first ventral is always wider than long and usually similar to the next ventral; small extranumerary scales sandwiched between posterior chin shields were not included); subcaudals were counted from the first adjoining pair behind the vent up to the pair in contact with the terminal spine (spine excluded); longitudinal dorsal scale rows were counted at or near midbody (halfway between tip of snout and vent), usually by starting the count caudad and reversing in the same way at the dorsal ridge (vertebral area), but taking special care to follow the inclination of the lateral rows (usually 3–4) of serrated and reduced scales; pattern cycles refers to the number of blotches or bands (‘saddles’) on the back, starting after the first (often chevron) marking on the nape (which often extends onto the back of the head) up to the marking above the vent (in cases of bilateral asymmetry, fusion of dorsal markings, or when these markings were unclear, lateral bars were used as guiding criteria); texture of the frontal scale was scored as indicated under ‘Character analysis’; depth of sulcus formed by the pair of prefrontal scales was scored as deep, moderate or weak (scales merely in contact) as illustrated by Gans (1959, Fig. 6). Anterior temporals are the (usually) elongated scales posterior to, and in contact with, the postoculars; the uppermost one is in contact with a parietal scale, while the lowermost one is in contact with a supralabial. Posterior temporals are in contact behind the anterior temporals; the uppermost in contact with a parietal and lowermost in contact with a supralabial. Occasional supranumerary scales between anterior and posterior temporals were excluded from counts. The temporal formula refers to the number of anterior temporals plus the number of posterior temporals (e.g. 2+3). Snout-vent length (SVL) (tip of snout to vent) and tail length (vent to tip of tail) were measured with the snake aligned against a millimetre rule, but SVL of curled specimens was measured using a length of string. Measurements of truncated tails were not included in calculations.
In some cases material reported on by Gans (1959) was also examined by the authors and our scale and size data were used (Appendix 1). Because of known differences in numbers of ventral and subcaudal scales between males and females (e.g. Gans 1959), specimens were sexed. Sex was determined by presence/absence of (one or both) inverted hemipenes (slit open to reveal spines if not obvious) in the tail base, presence of everted hemipenes, and/or presence of internal reproductive structures (eggs, ovaries, oviducts, testes, seminiferous tubules). Only sexed specimens were used for the analysis of numbers of ventral and subcaudal scales (except MSNG 6738 from Keren in Eritrea, one of only four specimens representing a new species, taken as female based on high ventral and low subcaudal counts; see below).
For the descriptive account of D. bazi five specimens from Egypt were examined, comprising three of the four specimens from this region reported on by Gans (1959, but excluding ZMB 5865), as well as a specimen (ZMB 51552) discovered by Barry Hughes among undetermined material at the museum in Berlin (Frank Tillack in litt. 9 April 2014), and a specimen (NMW 26879/1) in the Naturhistorisches Museum in Vienna. A set of colour photographs of the Berlin specimens, showing dorsal, ventral and lateral aspects of the body and tail, as well as close-up views of the head (including left and right sides), of these specimens, was also later consulted. For ZMB 5865, SVL and tail measurements, and number of pattern cycles on the back, follow Gans (1959), while numbers of midbody scale rows and pairs of subcaudals were determined by F. Tillack (these corresponded with Gans’ 1959 counts); other morphological data for this specimen was determined after examining the colour images. Data for specimen ZFMK 38415 (Erkowit, Sudan), which we refer to as Dasypeltis cf. bazi, was provided by W. Böhme, but colour photographs of this specimen were also examined. The ratio greatest frontal length/greatest width was determined after performing measurements on enlarged photographic images.
According to Nicola Maio (in litt. 8 September 2014) the two Egyptian specimens of Rachiodon scaber recorded by Panceri (1878), in the Comparative Anatomy section of Naples Museum, were transferred to the Museo Zoologico at the same university after the Second World War, but now appear to be lost. There is also no record of these specimens having been transferred to Museo Civico di Storia Naturale di Milano (S. Scali in litt. 10 September 2014). However, the old catalogue of the museum in Milan does record a few specimens from Egypt identified by Prof. Gasco in 1876, including one ‘Rachiodon scaber’ (catalogue number 1622 – photographic image of catalogue examined), but unfortunately none of these specimens still exist at the museum, possibly having been destroyed by fire in 1943 during the Second World War (S. Scali in litt. 26 September 2014). The Cairo specimen reported on by Jan (1863) is also no longer available in the Milan collection (S. Scali in litt. 26 September 2014).
The description of the new species from Somaliland and Djibouti (see below) was based mainly on data in Gans (1959) and details available from a set of high-resolution colour photographs of all proposed type specimens showing dorsal, ventral and lateral aspects of the body and tail, as well as close-up views of the head (including left and right sides), provided by the Natural History Museum (London) and Museum National d’Histoire Naturelle (Paris), although two proposed paratypes (BM 1992.501 and MNHN 84–707) were also examined by the first author while visiting these museums in September 2011 and October 2015, respectively. While the proposed holotype of the new species from Eritrea (see below) was examined by the first author at the natural history museum in Berlin, a set of close-up photographs was also supplied subsequently; a similar set of photographs of the proposed paratypes in Italian museums—which could not be examined personally—were also supplied, together with sex, SVL and tail length measurements, and ventral, subcaudal and midbody scale counts. For the new ‘5I’-patterned species from Arabia (see below), similar photos were provided of the proposed holotype (and one proposed paratype, BM 1987.1018) to allow for a detailed description, although these and other proposed types were also examined by both authors at the museum in London. Greatest frontal length/greatest width for most of the above specimens was determined by performing measurements on enlarged photographic images.
In addition to the data presented in this paper, comparative data consulted for the diagnoses of species (see below) were obtained from Duméril et al. (1854), Mertens (1954, 1955), Gans (1959, 1964), FitzSimons (1962), Broadley (1990), Trape & Mané (2006), Trape et al. (2012), Bates & Ineich (2012), Bates (2013) and Saleh & Sarhan (2016). In the case of D. atra, to avoid inadvertently including data for mainly brown or uniform phase specimens of other species of Dasypeltis, the only extralimital data used was for specimens from eastern Democratic Republic of the Congo (Mount Kabobo and the ‘districts’ of Oriental and Kivu) with ‘2M’, ‘2B’ and ‘2Bx’ dorsal colour patterns as identified by Gans (1959) in his appendix. An exceptionally low female subcaudal count of 50 for D. gansi (see Bates & Ineich 2012) was excluded from the ranges listed under ‘Diagnosis’ below, as were ventral and subcaudal counts (250 and 55 respectively) for a specimen of D. bazi identified as a male by Saleh & Sarhan (2016) but which is probably a female (see discussion below; these data were also excluded from the diagnostic key). Morphological data for D. parascabra were derived from the small type series together with a few additional specimens—for which data is available in Gans (1959)—referred to this species by Trape et al. (2012); ranges are as follows—males: ventrals 210–223, subcaudals 67–74; females: ventrals 224–237, subcaudals 57–64; both sexes: midbody scale rows 22–26, pattern cycles 56–70. Photographs of the holotype (SMF 46642) of D. s. loveridgei were available at: http://www.biologie.uni-ulm.de/cgi-bin/query_all/details.pl?id=107169&stufe=7&typ=ZOO&sid=T&lang=e (accessed 26 September 2014).
The following type specimens were examined for comparative purposes by the first author in September/October 2011: holotype (ZMB 1856) and two paratypes (ZMB 1857–8) of Dasypeltis scaber var. mossambicus Peters, 1864 (= D. scabra); holotypes of Coluber Palmarum (= D. palmarum, BM 1946.1.2.35), Dasypeltis inornatus (= D. inornata, BM 1946.1.2.47), Dasypeltis scaber var. breviceps Peters, 1864 (= D. scabra, ZMB 3886), Dasypeltis scaber var. fasciolata Peters, 1864 (= D. medici, ZMB 5737), D. lineolata Peters, 1878 (= D. atra, ZMB 9224, see below), D. confusa (MNHN 2006.0315) and D. sahelensis (MNHN 2006.0315); paratypes of D. medici lamuensis (= D. medici, see below, BM 1951.1.3.72–3, BM 98.1.8.15, BM 87.11.3.31), D. gansi (MNHN 2006.0308) and D. latericia (MNHN 2006.0314); and syntypes of Rachiodon abyssinus (= D. abyssina, see below, MNHN 6567, 7227) and Dasypeltis macrops Boulenger (= D. fasciata, BM 1946.1.5.17–8). Photographs of the syntypes (ZMB 22516A, ZMB 22516B , ZMB 27761) of D. scabra var. atra (= D. atra), provided by F. Tillack, were also examined.
Statistical and multivariate analyses.—Univariate analysis was conducted using the computer software package Statistica v. 6. Two kinds of ordination comparisons were conducted in order to determine whether samples were distinguishable using a multi-dimensional approach. For Principal Components Analysis (PCA), IBM SPSS Statistics v. 20 was used, while Statistica Six Sigma was used for Discriminant Function Analysis; PAleontological STatistics v. 3.08 was used for PERMANOVA and to produce graphs. As all population groups (= species) discussed below are considered real geographical entities, it was considered appropriate to conduct discriminant function analysis to determine if the groups are distinguishable. Because of known sexual dimorphism in numbers of ventral plates and pairs of subcaudal scales (Gans 1959; see below), males and females were analysed separately. Eleven meristic variables (characters as defined above and by Gans 1959, and discussed below) were used in the analyses: numbers of ventrals, subcaudals, midbody scale rows, supralabials (counts for left and right sides of the head combined), preoculars (combined count), postoculars (combined count), anterior temporals (combined count), posterior temporals (combined count) and pattern cycles; and the interval variables: frontal pitting (coded as 0 if smooth, 1 if present only as one or two rows at the edges, and 2 if extensive), and depth of the inter-prefrontal sulcus (0 if weakly marked, 1 if fair/medium, and 2 if deep). Only specimens with full sets of data were included (marked by an asterisk in Appendix 1). Analyses were conducted for the entire study area, but also separately for the Horn of Africa (Ethiopia, Eritrea, Djibouti, Somalia) + Egypt (excluding D. medici, represented by only a single Somalian specimen with a full set of data) to determine whether the new species were distinguishable from all other congeners in that region.
Species concept and species delimitation.—We apply a lineage-based species concept whereby a species is represented by an independently evolving metapopulation lineage (see Frost & Hillis 1990; De Queiroz 1998, 2007). Morphological characters were the operational criteria for species delimitation.
Broadley (1983, also 1990) relegated Dasypeltis scabra loveridgei Mertens, 1954 from Namibia to the synonymy of D. scabra, but the former has a distinctive dorsal colour pattern (hourglass-shaped dorsal saddles alternating with more-or-less square to rectangular lateral bars; ‘6N’ of Gans 1959) and almost always six supralabials (2nd and 3rd in contact with the orbit) on either side of the head (Mertens 1954; Bates et al. 2009; Bates et al. unpubl. data). Apart from Mertens’ (1954) types, Gans (1959) provided data for additional Namibian ‘6N’ specimens, all but a few having the 6[2,3] supralabial formula. This combination of characters is unique to D. s. loveridgei which we therefore consider a good evolutionary species and formerly elevate to species status as Dasypeltis loveridgei Mertens, 1954 comb. nov. It should be noted that one of Mertens’ (1954) paratypes has the ‘5N’ dorsal pattern and 7[3,4] supralabial formula and is referable to D. scabra (Gans 1959: 95, 219; Bates et al. in prep.).
Character analysis
- Colour phases.—Gans (1959, see his Figs 9 & 10) recognised 13 colour phases (dorsal colour patterns) for the genus Dasypeltis, with only ‘inornata’ (‘1’), ‘loveridgei’ (‘6’) and ‘palmarum’ (‘7’) not found in north-eastern Africa. The ‘montane’ form [= atra] included melanistic (‘2M’), uniform brown (‘2B’), and patterned (‘2Bx’, dorsal saddles often confluent, or in partial contact, with lateral markings) phases (Fig. 17A–M). Gans (1959) recorded the ‘lineolata’ (‘9’, dorsum dark with narrow, interrupted, pale transverse bands) phase [= D. atra] only from central Kenya (Fig. 17D), but some specimens from other areas have similar markings (see Fig. 17E, Ethiopia). ‘Dasypeltis scabra’ included true D. scabra (‘5N’, variable, but dorsal saddles not linked to lateral bars, see Gans 1959) (Figs 5A–H & 6); D. bazi (5N-like but with large, cranio-caudally elongate, brown saddles, usually bifurcated at each end with a fairly shallow but distinct notch, and spot or blotch-like lateral markings; Gans 1959, Pl. 6, Fig. 1) (Figs 7A, 9A, 11 & 12); D. taylori sp. nov. (‘5N’-like but with large, closely-spaced, square to slightly rectangular saddles often separated by narrow, sharply-defined white crossbars; Gans 1959, Pl. 6, Fig. 6) (Figs 13 & 14); D. confusa (‘5L’, saddles and lateral bars linked; Gans 1959, Pl. 8, Fig. 6) (Fig. 4A–D); and D. arabica sp. nov. (‘5I’, rectangular saddles and lateral bars enclose pale inter-saddle areas; Gans 1959, Pl. 6, Fig. 4) (Figs 27A & 29A). Dasypeltis fasciata (‘4’) was distinguished by its ‘chequerboard’ pattern being expressed only on the interstitial skin and lower edges of the scales (Gans 1959, Pl. 3, Figs 1–4) (Figs 19A & C). Dasypeltis medici, distinguished by the darkly pigmented edges of the paired apical pits on its dorsal scales, occurred as both patterned (‘8’, rectangular saddles with distinct white interspaces usually in contact with elongate lateral bars) and uniform or largely uniform but feebly marked (‘3’ – subspecies lamuensis) phases (Gans 1959, Pl. 13) (Fig. 2A–E). Dasypeltis crucifera sp. nov. has a unique dorsal pattern consisting of dark brown oval saddles, and elongated lateral bars, which together almost completely enclose pale cross-markings that form the inter-saddle areas (Figs 23A & 25A). Gans (1959) did not recognise D. abyssina, which is now known to occur in both (dorsally) patterned (‘5N’-like but with mostly oblique dark saddles, usually wider than long, and often confluent or forming zigzags on the back and tail) and uniform grey-brown phases (see below and Fig. 20A, C–G).
- Number of pattern cycles (Table 1).—This is a very variable character, but patterned atra, D. abyssina, D. arabica sp. nov., D. medici and D. fasciata usually have counts of 70 or above, and are usually distinguishable from D. bazi, D. taylori sp. nov., D. crucifera sp. nov., D. confusa and often also D. scabra in this regard. In the study area, high numbers of cycles (108–141) in D. fasciata distinguish it from all other species except D. atra and a couple of specimens of D. medici; while the lowest counts are for D. bazi (39-49, 38 in a specimen reported by Saleh & Sarhan 2016) and D. taylori sp. nov. (44-59).
INSERT TABLE 1
- Number of ventrals (Table 2).— arabica sp. nov. and D. abyssina have the highest counts, followed by D. fasciata and D. medici. Due to wide variation within species, and extensive overlap of ranges between species, this character on its own does not help to distinguish D. atra, D. bazi, D. taylori sp. nov., D. crucifera sp. nov., D. confusa and D. scabra. In D. medici, males and females often have similar ventral counts.
- Number of subcaudals (Table 2).—The two most arboreal species, fasciata and D. medici, have the highest subcaudal counts, averaging over 68 in both sexes. There is considerable geographical variation in D. scabra, with the highest counts in the south (Tanzania). Dasypeltis confusa, D. scabra, D. bazi (based only on specimens examined, i.e. excluding data from Saleh & Sarhan [2016] as some of their specimens may have been incorrectly sexed), D. taylori sp. nov., D. atra, D. abyssina, D. crucifera sp. nov. and D. arabica sp. nov. have similar mean counts (59–63 in males, 48–56 in females).
INSERT TABLE 2 (FULL PAGE)
- Type of head pitting with reference to the frontal shield.—Gans (1959) recognised three states: N = none, M = marginal pitting (his Fig. 2B), and A = entire surface pitted (his Fig. 2A). Pits may take the form of swollen pimple-like protrusions on the frontal, especially centrally. In the present study this scheme has been refined to show five states: 0 = smooth, no pitting; 1 = a single row of pits on the posterior and lateral margins of the frontal; 2 = more extensive marginal pitting (Fig. 2B of Gans 1959); 3 = extensive pitting, but with centre of frontal still smooth (Fig. 2A of Gans 1959); 4 = entire frontal pitted and centre of shield often rugose or corrugated. In the study area Dasypeltis medici and taylori sp. nov. almost always have extensively pitted frontals (States 3 or 4), while most other species usually have marginal pitting (States 1–2), but in D. scabra and D. bazi the pitting varies from absent (State 0) to extensive (State 4). The frontal shields of five specimens of D. bazi illustrated in Saleh & Sarhan (2016, photographs in Fig. 5) appear to be extensively pitted and sometimes also corrugated (States 3 or 4).
- Depth of inter-prefrontal sulcus.—This character was classified in the same way as by Gans (1959). The sulcus was moderately to deeply sunken in taylori sp. nov., D. arabica sp. nov. and some populations of D. scabra (but usually moderately sunken to absent elsewhere), whereas it was usually weakly marked (scales merely in contact) or moderately developed in all other species.
- Number of supralabial shields.—In north-eastern Africa and Arabia there are normally seven supralabials, the third and fourth entering the orbit. The 6[2,3] formula (meaning six supralabials on either side of the head, with the 2nd and 3rd in contact with the orbital) is very rare except in loveridgei comb. nov. and a few other populations of the D. scabra species complex in southern Africa (Mertens 1954; Bates & Broadley in prep.). Occasional variations (on one or both sides of the head) in the study area were: 5[2,3], 5[3], 6[2,3], 6[3], 6[3,4], 7[2,3,4], 7[3,4,5], 8[3,4], 8[3,4,5] and 8[4,5].
- Number of postoculars.—In the study area most specimens have a single preocular and two postoculars on either side of the head, but a single postocular is commonly found in western populations of atra (e.g. 93.0% of Rwanda/Burundi snakes [N = 43] had a single postocular on either side; by contrast 75.0% of Ethiopian snakes [N = 28] had two postoculars on either side). The occurrence of a single postocular is also high in extralimital populations―in a sample of 126 specimens of D. atra from eastern Democratic Republic of the Congo, examined by the first author at the Royal Museum for Central Africa (Tervuren, Belgium), as many as 73.8% had one postocular on either side of the head, 14.3% had one postocular on one side and two on the other, while only 11.9% had two postoculars on both sides of the head.
- Number of anterior and posterior temporals.—A temporal formula of 2+3 was common for all species, with 2+2 and 2+4 occuring frequently. Two anterior temporals on either side of the head was by far the most common condition for most species in the study area; as was three posterior temporals on either side, although counts of two or four were common. Although Saleh & Sarhan (2016) also gave this formula as 2+3 for bazi, in the single Fayoum specimen examined for this study, one side of the head had three anterior temporals, while another Egyptian specimen had a formula of 3+3 on both sides of the head.
- Number of dorsal scale rows at midbody.—There was considerable overlap in numbers of midbody scale rows between the different species. However, northern species ( arabica sp. nov., D. abyssina, D. crucifera sp. nov., D. bazi and D. taylori sp. nov.) never had more than 25 rows (21–25 versus 22–28 rows in other species). The highest means (≥25) were for D. scabra and D. confusa, and the lowest (≤23.5) were for D. abyssina, D. bazi (based only on specimens examined, i.e. excluding data from Saleh & Sarhan [2016], see below), D. crucifera sp. nov. and D. fasciata.
- Relative tail length (Fig. 1).—Although there was much overlap in this character for most species (see species accounts), including those for which reasonable samples were available as shown in the figure below, Dasypeltis medici males and females usually had distinctly longer tails relative to SVL compared to males and females, respectively, of other species, while fasciata tended to have a slightly longer tail than the other species. Figure 1 also illustrates sexual dimorphism in tail length, with males usually having longer tails relative to SVL; as well as sexual dimorphism in body size, with the largest females exceeding the size of the largest males.
INSERT FIG. 1
Systematics
Apart from the species discussed and compared below, two available names currently in the synonomy of D. scabra (see Gans 1959; Broadley 1990) deserve mention in light of the fact that material from three populations currently identifiable as, or referable to, D. scabra are here to be described as new species. Dasypeltis scabra mossambica (Tete and Boror in Mozambique) and Dasypeltis scabra breviceps (‘Cafferland’, i.e. Eastern Cape Province, South Africa) were described by Peters (1864). The three syntypes (ZMB 1856, juvenile male; paratypes 1857 and 1858, both adult females) of the former and holotype (ZMB 3886, adult female) of the latter were examined for this study. The types of D. s. mossambica have ‘5N’-type dorsal patterns with moderate-sized saddles generally round to oval in shape, nasal divided below the nostril, supralabial formula 7[3,4] on either side of head, postoculars two on either side, frontal smooth in juvenile but fully pitted (State 4) in adults. For other characters: ZMB 1856, inter-prefrontal sulcus weakly marked, ventrals 207, subcaudals 67, midbody scale rows 25, pattern cycles 57; ZMB 1857: sulcus deeply marked, 236, 60, 25, 50; ZMB 1858: sulcus moderately marked, 237, 59, 27, 61. The holotype of D. scabra breviceps has a ‘5N’-type dorsal pattern but the saddles and lateral bars are somewhat narrow and irregular, nasal divided below the nostril, supralabial formula 7[3,4] on either side of head, postoculars 2 on either side, frontal fully pitted (State 4), inter-prefrontal sulcus deeply marked, ventrals 203, subcaudals 61, midbody scale rows 27, pattern cycles 63. Peters (1864) noted that the ground colour of D. scabra mossambica was yellowish. Both of the above names are clearly synonyms of D. scabra.
Squamata: Colubridae: Boigini: Dasypeltis Wagler, 1830
Dasypeltis medici (Bianconi)
Eastern Forest Egg-eater
Fig. 2A–F
Dipsas medici Bianconi, 1859: 277. Type locality: ‘Mozambique’ [= Inhambane].
Dasypeltis abyssina ? (not Duméril, Bibron & Duméril) Stejneger, 1893: 733 (Mt Kilimanjaro, Tanzania).
Dasypeltis scabra var. F Boulenger, 1894: 356 (part, Mt Kilimanjaro).
Dasypeltis scaber (not Linnaeus) Loveridge, 1929: 28 (part) & 1936: 256 (Mt Mbololo, Lamu Island); Uthmöller, 1937: 356 (part, Gomberi, Mt Kilimanjaro); Scortecci, 1939: 276 (part, Belet Amin, Somalia).
Dasypeltis medici medici Gans, 1957: 3 & 1959: 157; Malonza et al. 2011: 160 (Kitobo Forest, Kenya); Göthel, 2015a: 19 & 2015h: 89.
Dasypeltis medici lamuensis Gans, 1957: 5. Type locality: Lamu Island, Kenya, holotype MCZ 40581; Loveridge, 1957: 289 (check-list); Gans, 1959: 159 & 1961: 74; Spawls, 1978: 10; Lanza, 1983: 223 (Alessandra Island near Gelib, zone of Mareri SW of Gelib, Belet Amin); Broadley & Howell, 1991: 32; Spawls et al., 2002: 416; Spawls et al., 2006: 129; Largen & Spawls, 2010: 490; Göthel, 2015a: 19, 2015c: 30, 2015d: 50, 2015f: 65 & 2015h: 89.
Dasypeltis medici Gans, 1971: 461; Largen & Parker, 2004: 26; Branch, 2005: 59 & 2014: 68; Vets & Safari, 2014: 43 (University of Dodoma).
Description. Dorsal colour pattern varies considerably. Patterned specimens may be similar to D. scabra: grey to tan with narrow, darker, rectangular saddles (longer than wide) separated by narrow cream to white interspaces, with irregular narrow dark lateral bars situated for the most part adjacent to the pale interspaces. The saddles may be much elongated and closely set and give the appearance of a broad dark vertebral stripe broken by more-or-less equally spaced pale flecks. Pattern cycles 57–108 (mean 74.0, N = 24); 1–5 chevrons (pointing forward) on nape and anterior part of body (N = 13). Some specimens are uniform pink, orange, red, grey, fawn or brown above (Spawls et al. 2002), but specimens with intermediate patterns also occur, e.g. reddish (Fig. 2C–D) or grey (Fig. 2E–F) with fairly vague dark dorsal markings and mere indications of whitish interspaces (e.g. CAS 135751, FMNH 251325, ZMUC 6111); NML 1699 is reddish-brown with 3–4 vague chevrons only (anterior part of body), and a cream venter. Edges of apical pits on dorsal scales darkly pigmented. Venter usually cream, often with fine grey stippling (may be extensive as in NMW 26926); but occasionally yellowish-cream (e.g. NMK S 3956, Mount Warges; ZFMK 63582, near Mombasa [with extensive grey speckling]); and salmon-coloured with central part of each plate dark brown anteriorly in FMNH 142635 (vicinity of Nairobi).
Preoculars usually one on either side of head, but one on one side and two on the other in NMW 9964/18 and ZMB 8597, and two on either side in ZMUC 6143 (N = 61); postoculars usually two on either side of head, but one in 10.3% of specimens, and one on one side and two on the other in 4.6% of specimens (N = 87); frontal shield usually extensively pitted (States 3–4), but marginally pitted (States 1–2) in 10.5% of specimens (including three snakes ≥422 mm SVL) and smooth (State 0) in one adult (MSNG 42710b) (N = 57); inter-prefrontal suture usually weakly marked, but moderately marked in 18.9% of specimens, and deeply marked in AMNH 143717 (N = 53); supralabials 12–15, usually 14 (both sides of head), usually 7 [3rd and 4th entering orbit] on either side of head, but variations are: 6[2,3] on both sides in NMK 1699, 6 [2,3] on right side in MSNG 427210b and USNM 16755, 6[3,4] on both sides in NMK 2535 & 3249, 7 [2,3,4] on right side in ZMUC 6124, 8 [3,4] on left side in SMNS 4216A, and 8[3,4,5] on left and 7 [3,4,5] on right side in SMNS 4216F (N = 85); anterior temporals usually two on either side of head (70.5%), occasionally three (14.8%), 3 left:2 right or 2:3 (13.1%) and 4:3 in USNM 16755 (N = 61); posterior temporals usually three on either side of head (60.7%), occasionally 3:4 or 4:3 (14.8%), four (14.8%), 3:2 or 2:3 (4.9%), 5:4 in MCZ 40580, five in ZMB 17453 (N = 61), and two (left side, right damaged) in ZMB 48150; temporal formula on either side of head usually 2+3, occasionally 2+4 or 3+3; midbody scale rows 22–27 (mean 24.8 ±0.97, N = 57); ventrals 220–252 in males (N = 29), 218–252 in females (N = 49); subcaudals 75–109 in males (N = 27), 61–90 in females (N = 47). Ratio SVL: tail length 3.5–4.7 in males (N = 27), 4.0–6.1 in females (N = 44); total length: tail length 4.5–5.7 in males (N = 27), 5.0–7.1 in females (N = 44).
INSERT FIG. 2
Size. Largest male (NMK 2631 – Gede, Kenya) 679 mm snout-vent length [SVL] + 146 mm tail length = 825 mm total length; largest female (FMNH 142635 – ‘vicinity of Nairobi’, Kenya) 912 + 149 = 1061 mm.
Habitat. Coastal and montane forest and savannah at elevations from sea level to about 2500 m on Mount Warges (Kenya). In southern Africa it occurs in lowland evergreen forest (Broadley 1990).
Distribution. Widely distributed in the eastern parts of sub-Saharan Africa, from Mareri in southern Somalia southwards through eastern and central Kenya, eastern Tanzania, Malawi, extreme eastern Zimbabwe, Mozambique, and KwaZulu-Natal in South Africa as far south as St Lucia (see Broadley 1990; Spawls et al. 2002; Bates et al. 2014; Fig. 3). There is also an isolated Zambian record (AJL 3027) from riparian forest on the Mululangana River in the Zambezi Valley, 26 km SW of Chirundu [SE 1628B1] (Broadley et al. 2003).
Sympatry. Sympatric with D. scabra at Belet Amin (Somalia), Kilifi (Kenya), and Amani, Dodoma, Morogoro and Zanzibar (Tanzania); sympatric with D. atra at Nyambene Hills (Kenya) and Moshi (Tanzania); sympatric with both of these species at Mt Kilimanjaro in Tanzania (Appendix 1).
Localities. SOMALIA. Belet Amin [NE 0042B2] MSNG 42710b; near Mareri (Labadaah to Bula Yaag) [NE 0042D1] CAS 148077, MZUF 27155. KENYA. Chyulu Hills [SE 0037D2] NMK 402; Diani Beach, Ukanda [SE 0439B3] Fig. 2E–F; Gede [SE 0340A3] AMNH 88631, NMK 2597, 2619, 2631, 3200; Giriama [SE 0339D3] NMK 480; Kikambala [SE 0339D4] NMK 1050; Kilifi [SE 0339D2] BM 51.1.3.72–3; Lamu Island [SE 0240B4] MCZ 40581–2 [holotype and allotype of D. medici lamuensis]; Malindi [SE 0340A1] NMK 1561, 2522, 2526–7, 2529–33, 2535, 2538, 2587, 3290, 3292, ZMB 50191; Malindi, Sabaki River [SE 0340A1] photographic record (dorsum and top of head plain pale orange, edges of apical pits on dorsal scales darkly pigmented) and bottom photograph in Spawls et al. 2002: 417; near Mombasa [SE 0439B1] ZFMK 63582; Mombasa, 30 km south of [SE 0439B3] NMW 26926/1; Mount Mbololo [SE 0338A4] MCZ 40580; Maungu, Teita [SE 0338D2] BM 98.1.8.15; Mount Warges [NE 0037C4] NMK 3956; ‘vicinity of Nairobi’ [SE 0136B4] FMNH 142635; Nguni, Mwingi [SE 0137D2] NMK 2377; Nyambene Hills, Meru [NE 0037B2] NMK 1699; Taita [SE 0338A4] ZMB 9244; Takaungu [SE 0339D2] ZMB 17496; Tana River, Mchelelo Forest [SE 0140A3] NMK 4131; Taveta, Kitobo Forest [SE 0337B3] NMK 4121; Tiwi, Kwale [SE 0439B1] CAS 135751; Voi to Ndi [SE 0338D3] BM 98.1.8.14; Watamu [SE 0340A3] (Fig. 2C). TANZANIA. Amani [SE 0538B1] MCZ 23274, ZMB 37598, ZMUC 6124; Bombani, East Usambara Mtns [SE 0538B1] ZMUC 6123, 6133; Dar-es-Salaam [SE 0639C4] NMW 9964/18; Dar-es-Salaam, vicinity of [SE 0639C4] (Fig. 2A); Dodoma, University of Dodoma campus [SE 0635B1] photographic record in Vats & Safari (2014); East Usambara Mtns (Fig. 2B); FMNH 251325, ZMUC 6143; Gomberi, Mount Kilimanjaro [SE 0337A2] (Uthmöller 1934); Kibognoto [SE0337 A1] ZMB 37600; Kimboza Forest [SE 0737B2] ZMB 48150; Longido [SE 0236D1] ZMUC 6144; Mafia Island [SE 0739D4] MCZ 54827, NMZB 11265, SMF 19460; Makindu River [SE 0738C3] MCZ 18206; Mlinga Forest Reserve [SE 0538B1] ZMB 17318; Morogoro [SE 0637D3] AMNH 143717, USNM 62901, 62904, 62906, ZMB 27616; Moshi [SE 0337A4] SMNS 4216A–F; Mount Kilimanjaro [SE 0337A2] BM 87.11.3.31, USNM 16755; Mount Kwaraha [SE 0435B2] ZFMK 82342; Msolwa, Rubeho Mtns [SE 0736B4] ZMUC 6121; Nyange, Uluguru Mtns [SE 0637D4] MCZ 23273; Tanga [SE 0539A1] Fig. 2D and middle photograph of reddish-brown, weakly marked specimen in Spawls et al. 2002: 417; Usa River near Arusha [SE 0336B3] CM 36586; Usambara Mountains, ZMB 17453, and Spawls et al. 2002: 417; Zanzibar Island [SE 0639A2] BM 50.1.5.40, NMK 3249, ZMB 5737, 17219, ZMUC 6111; Zanzibar Coast, ZMB 8597.
Conservation. In terms of the regional (study area) and global distribution of this species, using IUCN (2012, 2016) criteria, it appears to be widespread and abundant and should be regarded as Least Concern.
Remarks. The northern-most populations of D. medici were separated as a subspecies, D. medici lamuensis, by Gans (1957). This taxon was diagnosed by the lack of the typical medici colour pattern (saddles and lateral bars) on the body (although this may be present but poorly defined, and Gans [1964] later noted that a faint typical medici pattern may be present in juveniles) and lower numbers of ventrals in both sexes. However, medici (marked), uniform, and ‘intermediate’ patterns are found in many specimens from south-eastern Kenya, especially at Malindi and Gede (AMNH, NMK and ZMB material not examined by Gans; Appendix 1: Malindi: in alcohol usually uniform and mostly pale brown [e.g. NMK 1561, 2619], occasionally grey [e.g. NMK 2532], but two specimens [NMK 2526, grey; NMK 2530, pale brown] have very weakly marked darker saddles, while two young snakes [NMK 2540, 422 mm SVL; ZMB 50191, 461 mm SVL] have the typically marked [but not bold] medici pattern; Gede: uniform pale brown, but NMK 2631 is grey with very vague indications of darker vertebral blotches and whitish interspaces). Reddish, largely uniform specimens, with faint vertebral blotches and whitish interspaces—from Watamu (SE Kenya) and Tanga (N Tanzania)—and a greyish, largely uniform specimen, with similar markings—from Diani Beach (SE Kenya)—are illustrated in Fig. 2. Weakly marked specimens with indications of dorsal and lateral markings are also occasionally found further south, e.g. Mlola Forest on Mafia Island in Tanzania (NMZB 11265) and Bazaruto Island off the southern coast of Mozambique (NMZB 12037). The average ventral counts are indeed lower in northern populations (Somalia + Kenya), but there is considerable overlap with typical southern populations (Appendix 1), and ventral counts are actually lowest in the southern-most population in KwaZulu-Natal, South Africa: 206–215 in three males (TMP 28779, 46041, 56547), 214–223 in three females (TMP 44652, 47840, 78994) (D.G. Broadley unpubl. data). In light of this variation in colour pattern and ventral counts, we here relegate Dasypeltis medici lamuensis Gans, 1957 to the synonomy of Dasypeltis medici (Bianconi, 1859).
INSERT FIG. 3 (FULL PAGE)
Dasypeltis confusa Trape & Mané
Diamond-back Egg-eater
Fig. 4A–D
Dasypeltis scabra scabra (not Linnaeus) Loveridge, 1955: 46 (South Sudan).
Dasypeltis scabra (not Linnaeus) Gans, 1959: 141 (part, 5L specimens from South Sudan and Uganda); Pitman 1974: 116 (part, Uganda); Hughes, 1997: 68 (part, Uganda); Behangana & Goodman, 2002: 66 (part, Uganda).
Dasypeltis confusa Trape & Mané, 2006: 28. Type locality: Ibel, Senegal, holotype MNHN 2006.0303; Göthel 2015h: 79.
Description. Dorsum grey with darker markings, usually in the form of rhombic saddles (often diamond-shaped with sharpest points aligned antero-posteriorly) which are confluent with the lateral bars (i.e. the ‘5L’ linked pattern of Gans 1959, Pl. VIII, Fig. 6); dark markings usually have a cream to white margin around them, emphasising the pattern; pattern cycles 50–79 (mean 61.7, N = 30); dorsal saddles are not always joined to the lateral bars, so that the dorsal pattern may be similar to that of D. scabra (‘5N’) over small portions of the back in some specimens. A few specimens from South Sudan (six of the 14 from Torit – see Gans 1959: 203) and Uganda (MCZ 47801, 47830, UMMZ 88530) are plain brown dorsally. Venter cream-white, often with grey stippling at the edges.
Preoculars one on either side of head (N = 40); postoculars usually two on either side of head, but only one on left side in FMNH 62216 (N = 45); frontal shield for the most part smooth, usually with 1–2 rows of shallow pits at the edges (States 1–2), but occasionally completely smooth and without pits (State 0) (ZMB 56436, UMMZ 88529) (N = 38); inter-prefrontal suture usually weakly marked, but moderately marked in 21.7% of specimens, and well marked (deep) in BM 1954.1.12.9 (N = 23); supralabials 11–14, usually 14 (both sides of head), usually 7 [3rd and 4th entering orbit] on either side of head, but variations are: 6 [2,3] (BM 1954.1.12.8), 6 [3,4] on left side of head (FMNH 58336), and 6 [3,4] on left and 5 [3] on right (ZMB 77441) (N = 42); anterior temporals usually two on either side of head (95.3%), but 1 left: 2 right in BM 1954.1.12.8 and 2: 1 in BM 1954.1.12.9 (N = 43); posterior temporals usually three on either side of head (62.8%), two in NMW 9964/7, occasionally 3:4 or 4:3 (11.6%), four (11.6%), 2:3 or 3:2 (9.3%), and five in MCZ 47815 (N = 43); temporal formula on either side of head usually 2+3, occasionally 2+4 or 2+2; midbody scale rows 23–27 (mean 25.0 ±1.03, N = 40); ventrals 199–224 in males (N = 14), 212–239 in females (N = 30); subcaudals 51–67 in males (N = 12), 48–61 in females (N = 24). Ratio SVL: tail length 4.7–5.8 in males (N = 12), 4.9–7.5 in females (N = 20); total length: tail length 5.7–6.8 in males (N = 12), 5.9–8.5 in females (N = 20).
INSERT FIG. 4
Size. Largest male (NMK 489 – Kacheliba, Kenya) 424 + 74 = 498 mm; largest female (FMNH 58496 – Juba, South Sudan) 575 + 92 = 667 mm.
Habitat. Found in savannah at elevations of 500 m to 1200 m.
Distribution. From Senegal to Nigeria in West Africa, and from Cameroon south to Angola and north-western Zambia, then eastwards through the Central African Republic, northern Democratic Republic of the Congo (‘5L’ patterned specimens in Gans 1959; Trape & Mané 2006; Trape et al. 2012; Bates 2013; Broadley & Bates unpubl. data), southern South Sudan, western and northern Uganda, to the Rift Valley at Kacheliba and Mumias in western Kenya, with an isolated record at Lake Kivu in western Rwanda (Fig. 3). It may also extend into south-western Ethiopia, based on two specimens with imprecise/questionable locality data (see below), but this requires confirmation.
Sympatry. Sympatric with D. atra 10 km S of Juba (South Sudan), Busingiro and Gulu (Uganda) and Kacheliba (Kenya); sympatric with D. scabra at Lake Kivu (Rwanda). (Appendix 1).
Mimicry. The dorsal pattern of D. confusa closely resembles that of the night adders Causus rhombeatus (Lichtenstein), C. bilineatus Boulenger, C. defilippii (Jan) and C. maculatus (Hallowell) which all occur in north-eastern Africa (Spawls et al. 2002; Rasmussen 2005; Dobiey & Vogel 2007).
Localities. SOUTH SUDAN. Juba [NE 0431D3] FMNH 58496, ZFMK 29694; Logutoic, MCZ 53401; Lokwi [NE 0432A2] FMNH 62216; Mongalla [NE 0531D1] MCZ 53402; Nimule [NE 0332C1] MCZ 53403; ‘Sudan’ (probably East Equatoria state, or vicinity of Wadelai, Uganda; see Remarks under D. scabra) NMW 9964/7; Torit [NE 0432B1] FMNH 58361–3, 62207–14, 62217, MCZ 53404–5; Yei [NE 0430B1] FMNH 58336. ETHIOPIA. No precise locality, NMW 9964.2; ‘Addis Ababa’, ZMB 56436. UGANDA. Ajai Game Reserve [NE 0231C] MUZM —; Bisu, Kigezi [NE 0131C] BM 51.1.3.69; Bugoma Forest [NE 0130B4] LACM 39043; Busingiro, Budongo Forest [NE 0131D3] MCZ 47815; Gulu, Acholi [NE 0232C4] MCZ 47830; Kyambura Game Reserve [SE 0030A1] NMZB 14333, 15411; Lira, Lango [NE 0232B4] BM 51.1.5.39, MCZ 47801, UMMZ 88529–30; Moyo, West Madi [NE 0331D1] BM 51.1.3.84; Nakosa [SE 0030A3] LIV 1960.51.2; Nyanshimbo [SE 0030A1] LIV 1962.106.19; Sana [SE 0030B4] LIV 1960.67.2; Semliki [NE 0130A2] BM 1954.1.12.8 (‘120’) & 1954.1.12.9 (‘121’); Serere, Teso [NE 0133C2] BM 1976.848. KENYA. Kacheliba [NE 0135A3] NMK 485, 489; Mumias [NE 0034A4] CAS 141784. RWANDA. Lake Kivu [SE 0229A1] ZMB 77441.
Conservation. In terms of the regional (study area) and global distribution of this species, using IUCN (2012, 2016) criteria, it appears to be widespread and should be regarded as Least Concern.
Remarks. The plain phase specimens from South Sudan and Uganda reported on here (see above) are the first of their kind to be documented. Dorsally uniform, or mostly unpatterned, specimens (brown or grey) are also known for D. scabra (southern Africa: e.g. De Waal 1978; Broadley 1990), D. medici (Gans 1959; Spawls et al. 2002; this study), D. gansi (Trape & Mané 2006; Bates & Ineich 2012), D. atra (Gans 1959, 1964; Spawls et al. 2002) and D. abyssina (see below).
Dasypeltis scabra (Linnaeus)
Common Egg-eater
Figs 5A–H & 6
Coluber scaber Linnaeus, 1758: 223. Type locality: ‘in Indiis’ [= “via the Cape … read as South Africa”, Flower 1933: 818; restricted to “probably near Cape Town”, Mertens 1954: 214].
Rachiodon abyssinus (not Duméril, Bibron & Duméril) Duméril, Bibron & Duméril, 1854: Vol. 7: 496 (part, “Nil Blanc”) (MNHN 7227, proposed paralectotype of R. abyssinus, see below).
Dasypeltis scaber, var. abyssinicus (lapsus) (not Duméril, Bibron & Duméril) Peters, 1864: 645 (Sennar, Sudan).
Dasypeltis scabra var. B Boulenger, 1894: 356.
Dasypeltis scabra Günther, 1894: 87 (E. Kikuyu, Kenya); Boulenger, 1896: 20 (between Comia & Mategoi, Somalia), 1897: 229 (Somalia) & 1898: 720 (Lugh, Somalia); Lönnberg & Andersson, 1913: 4 (Kismayu); Loveridge, 1929: 28 (part); Corkill, 1935: 18 (Blue Nile Province, Sudan); Loveridge, 1957: 288 (check-list); Gans, 1959: 141 (part), 1961: 73, 1964: 283, 1971: 461 & 1974: 34; Gans & Laurent, 1965: 67 (Mogadiscio); Lanza, 1972: 177 (Ilala Sala, Awash National Park), 1983: 223 (part) & 1990: 436 (part); Pitman, 1974: 116 (part); Spawls, 1978: 10 (part); Emmrich, 1985: 66, 68 (Oukam, Lake Zwai); Hoevers & Johnson, 1982: 199 (Mareri); Broadley & Howell, 1991: 32; Meirte, 1992: 57; Largen & Rasmussen, 1993: 340 (part); Hughes, 1997: 68 (part); Largen 1997: 89 (part); Behangana & Goodman, 2002: 66 (part, Uganda); Spawls et al., 2002: 418; Branch, 2005: 59 & 2015: 67; Spawls et al., 2006: 130; Ziliani et al., 2006: 193 (Mount Kulal, Kenya); Largen & Spawls, 2010: 490 (part); Geniez, 2015: 211 (Mount Kulal); Spawls, 2017: 31 (Maasai Mara).
Dasypeltis scaber Calabresi, 1927: 54 (Somalia); Scortecci, 1931: 208 (Giohar), 1939: 276 (part, Giohar, Belet Amin) & 1943: 285 (El Dire).
Dasypeltis scabra scabra De Witte, 1962: 139; Bourgeois, 1968: 284.
Dasypeltis scaber scaber Laurent, 1956: 258 (part).
Dasypeltis “scabra” Göthel, 2015a: 19 (part) & 2015h: 86 (part).
Dasypeltis cf. scabra Sindaco et al., 2013: 331 (part, Kenya).
Description. In the study area the dorsal pattern may vary quite considerably (see Gans 1959). Dorsum grey or light brown with darker markings (‘5N’ pattern of Gans 1959); saddles vary from rectangular to squarish, oval, subcircular, weakly hourglass-shaped, weakly transversely enlarged, or irregular, and often a mixture of two or more of these shapes (Figs 5 & 6). Dorsal saddles (46–94, mean 67.4, N = 107) are dark brown or black, and a few may be linked to the dark lateral bars. The centre of the saddle may be paler than the outer rim, as in MHNP 7227 (Sudan) where it is grey with a black edge (Fig. 6). Venter cream-white, usually with grey stipples, or dark blotches, at the edges; sometimes with large bold cones or blotches at the edges and/or mid-ventrally (see Gans 1959, e.g. Pl. 9). Top of head often with 1–2 anterior-pointing (sometimes fragmentary) chevrons and/or other dark markings.
Preoculars usually one on either side of head, but one on one side and two on the other in MCZ 30208 & USNM 62902, and two on either side in BM 01.1.3.4 & MSNG 28808 (N = 104); postoculars usually two on either side of head, but one on one side and two on the other in 6.0% of specimens, one on either side in 6.0% of specimens, and absent on both sides in ZMB 26832 (N = 133); frontal shield heavily pitted (States 3 or 4) in 57.7% of specimens (73.3% of specimens from Tanzania in the south of the study area, N = 30), marginally pitted (States 1 or 2) in 36.1% of specimens, and smooth (State 0) in 6.2% of specimens (N = 97); inter-prefrontal suture weak in 48.9% of specimens, moderately marked in 38.0% of specimens, and deeply sunken in 13.0% of specimens (N = 92); supralabials 11–16, usually 14 (both sides of head), usually 7 [3rd and 4th entering orbit] on either side of head, but variations are: 5[2,3] on right and 6[2,3] on left in ZMUC 6136, 6[2,3] on both sides in BM 68.2.29.101 and BM 1964.1069, 6[3,4] on both sides in ZMB 22518, 6[2,3] on one side and 7[3,4] on the other in FMNH 134939, MSNG 29239 and MZUF 2802, 6[3,4] on left and 7[3,4] on right in BM 68.2.29.100, 7[2,3] on both sides in NMK 3184, and 8[4,5] on both sides in ZMB 51835 (N = 123); anterior temporals usually two on either side of head (88.5%), occasionally 2 left:3 right or 3:2 (3.5%), three (3.5%), 2:1 (3.5%), and one in MRAC 18563 (N = 113); posterior temporals usually three on either side of head (64.6%), occasionally 3:4 or 4:3 (16.8%), 3:2 or 2:3 (7.1%), four (6.2%), two (4.4%), and five in PEM 5626 (N = 113); temporal formula on either side of head usually 2+3, occasionally 2+4 or 2+2; midbody scale rows 22–28 (mean 25.0 ±1.38, N = 102); ventrals 185–226 in males (N = 50), 202–249 in females (N = 76); subcaudals 47-73 in males (N = 45), 43–65 in females (N = 72). Ratio SVL: tail length 3.9–6.2 in males (N = 45), 5.3–8.9 (N = 65) in females; total length: tail length 4.9–7.2 in males (N = 45), 6.3–9.9 in females (N = 65).
INSERT FIGS 5 + 6
Size. Largest male (NMK 2965 – Kora, Kenya) 456 + 88 = 544 mm; LIV 1962.168.14 (Ruzinga, Ankole, Kenya) has a smaller SVL but is the longest male: 453 + 98 = 551 mm; largest female (NMK 3184 – Lake Naivasha, Kenya) 867 + 95 = 962 mm.
Habitat. Found in savannah and grassland from sea level to over 1500 m.
Distribution. From the south-western Cape of South Africa through Namibia, Botswana, Zambia, southern and western Democratic Republic of the Congo, much of south-eastern and eastern Africa as far north as Gojam area (too vague to map) in north-central Ethiopia, with isolated populations along the Nile Valley (see comments under D. bazi) in south-eastern Sudan and adjacent north-eastern South Sudan (see Gans 1959; Broadley 1990; Spawls et al. 2002; Fig. 3), as well as 50 km north of Kutum in western Sudan (see Remarks below).
Sympatry. Sympatric with with D. medici at Belet Amin (Somalia), Kilifi (Kenya), and Amani, Dodoma, Morogoro and Zanzibar (Tanzania); sympatric with D. confusa at Lake Kivu (Rwanda); sympatric with D. atra at Aware Melca and Lake Abiata (Ethiopia), Mulema (Uganda), Lake Naivasha and Mount Kulal (Kenya), Astrida (Rwanda) and Minziro Forest Reserve (Tanzania); sympatric with both D. medici and D. atra at Mt Kilimanjaro (Appendix 1).
Localities. ‘ARABIA’. No specific locality, MNHN 6565. SUDAN. Nil Blanc, MNHN 7227 (Fig. 5, proposed paralectotype of D. abyssina, see below); 40 km E of Rabak, near Kosti [NE 1333A1] RMNH 24399; Sennar State [NE 1334C], BM 09.10.15.35. SOUTH SUDAN. Melut [NE 1032A3] FMNH 134939; ‘Sudan’ (probably East Equatoria state, or vicinity of Wadelai, Uganda; see Remarks), NMW 9964/6. ETHIOPIA. No locality, NMW 15298; Aware Melca [NE 0939B2] BM 16.6.24.11–12; Arba Minch [NE 0637B1] AAU 494, 679, BM 1969.288; Awash National Park [NE 0839D4] (Figs 5G&H); Awash National Park, 2 km NE of entrance to [NE 0839D4] PEM 8602–3; Bulcha [NE 0638A1] ZFMK 17763; Fitu, 135 km ESE of Neghelle [NE 0540B1] ‘5N’/‘2Bx’ dorsal pattern, photographic records; Gojam, BM 1964.1069; Lake Abiata [NE 0738D1] BM 1973.3259; Lake Chamo (west shore) [NE 0537D3] AAU 489; Meki–Dukem area [NE 0838B] ZMB 51835, 77053; Neghelle, 31 km ESE of [NE 0539B2] vertebral stripe over most of back, photographic records; west of Omo River [NE 0436C], BM 00.11.29.3. SOMALIA. environs of Afgoi [NE 0245A1] MZUF 1118, 1133; Afgoi, 6 km E of [NE 0245A1] MZUF 24179; Belet Amin [NE 0042B2] MSNG 42710A; between Comia and Matagoi [NE 0143C] MSNG 29239; Ceelcillan [= El Ellan, NE 0242D1] MZUF 31459; Dinsoor [NE 0242B4] MZUF 24674; Giohar [NE 0245D3] MSNM 785; near Mareri (Labadaah to Bula Yaag) [0042D1] MZUF 27156; Lugh (= Luuq) [NE 0342D1] BM 98.1.28.14, MSNG 28808; Lugh to Ganane [NE 0342D] MZUF 21968; Maheddei Uen [NE 0245D3] MZUF 21968; Mogadishu [NE 0242A2] MZUF 2802; Uagadi [NE 0144D1] MSNM 974. UGANDA. Kaburangiri [SE 0031D3] LIV 1962.106.6; Kakoba [SE 0030C1] LIV 1960.67.6; Kayanja, LIV 1962.330.22; Mbarara, Kasheri [SE 0030D1] BM 51.1.5.79, 1961.1804, LIV 1959.83.1; Mulema [SE 0031D1] BM 03.12.2.21; Namatyobola [SE 0031D1] LIV 1962.106.5; Ruburogoto [SE 0130B1] LICVM 1960.67.4, 1960.67.7, 1962.106.35; Rukoni [SE 0030C4] LIV 1962.106.20; Ruzinga, Isingiro [SE 0030D4] BM 1959.1.5.59, LIV 1962.168.14; Rwensama [SE 0029B4] LIV 1960.67.8; Wadelai [NE 0231C2] BM 07.12.1.19. KENYA. Aberdare Mtns [SE 0036B] USNM 42016; Buna, Wajir [NE 0239D3] NMK 2824; about 45 km W of Ewaro Ngiro, near Narok [SE 0135B3] PEM 5627; Gatab, Mount Kulal [NE 0236D2] MSNPV CR790–2; Gilgil [SE 0036C2] BM 01.1.3.3–4, and Fig. 5E; Hell’s Gate, Naivasha [SE 0036C4] CAS 147836; Isiolo [NE 0037B3] CAS 130784–5, NMK 561; Kilifi [SE 0339D2] BM 51.1.3.70; Kithu-Uri, East Kikuyu [SE 0037A4] BM 93.11.21.72; Kora [NE 0038B1] NMK 2965; Laini, Tana River [SE 0420C2] MCZ 40584; Lake Naivasha [SE 0036C4] NMK 1415, 1472, 2083, 2343, 2853, 3184, 3319, NMP 1260; Loita Hills [SE 0135D2] Fig. 5D; Mombosasa, Witu [SE 0240B3] MCZ 40583; Moshi [0337C2] (Gans 1964); Naivasha [SE 0036C2] CAS 147837–8; Ol Ari Nyiro Ranch [NE 0036C2] NMK 3103; Solio Ranch, Laikipia [SE 0036B4] NMK 3052; Sotik, Uaso Nyiro [SE 0035C1] USNM 41974; Tsavo East National Park [SE 0338B] NMK 3021; Uaso Nyiro Plains, MCZ 11501; Voi [SE 0338B3] NMK 1022; Wajir [NE 0140C3] CAS 130661, NMK 2262, Fig. 5C, and top photograph in Spawls et al. 2004: 418; Wema [SE 0240A1] MCZ 40585. RWANDA. Astrida [SE 0229D1] MRAC 18559; 15 km S of Butare [SE 0229D1] PEM 5624; Gabiro [SE 0130C2] IRSNB 5499, MRAC 9209, 9214; Lake Kivu [SE 0229A1] ZMB 22518; Nyagatare [SE 0130A4] PEM 5625. BURUNDI. Kabuyenge [SE 0329A4] MRAC 18563; Kahiro District [SE 0330C3] MRAC 18565; Mugera [SE 0329B4] MRAC 18564. TANZANIA. Amani [SE 0538A4] ZMB 21062; Chai Camp [SE 0336B4] AMNH 50791; Dar es Salaam [SE 0639C4] NMW 9964/17, ZMB 26832; Dodoma [SE 0635B1] (Fig. 5A); Kibwesa [SE 0629B4] ZMB 22754; Kikombo, Dodoma [0635B2] USNM 72476; Maji Malulu, Usandawi [SE 043B4] MCZ 30205; Mapenduzi, Dodoma [SE 0635B2] PEM 5626; Masiko ya Tembo, vicinity of Sanya Chini [SE 0337A3] (Fig. 5B); Massailand, ZMB 11934a,b, 14097; Minziro Forest Reserve [SE 0131B1] ZMUC 6136–7; Morogoro [SE 0637D3] USNM 62902–3, 62905; Mount Kilimanjaro [SE 0337A2] BM 87.11.3.30; Mpwapwa [SE 0636B3] ZMB 13304, 19669; Ngare, Nanyuki [SE 0235C1] FMNH 27786; Serondera, Serengeti National Park [SE 0234 C4] NMK 1887;Tanga [SE 0539A1] NMW 9964/9; Ukerewe Island [SE 0233A1] MCZ 30207–8; Wembere Steppe [SE 0434A] ZMB 16753; Zengeragusa, Mkalama [SE 0434B1] MCZ 18208.
Conservation. In terms of the regional (study area) and global distribution of this species, using IUCN (2012, 2016) criteria, it appears to be widespread and abundant and should be regarded as Least Concern.
Remarks. MNHN 6565 is a female from “Arabia” (without precise locality data) with a ‘5N’ dorsal pattern consisting of 55 fairly broad saddles (some ‘diabolo’-shaped), 244 ventrals and 54 subcaudals. Although its provenance is uncertain, we refer this specimen to D. scabra.
According to Gans (1959: 11, 205) specimen BM 07.12.1.19 was collected in ‘Wadelai’, north-western Uganda, by Emin Pasha (also known as Emin Bey, baptised E.C.O.T. Schnitzer [1840-1892]: Chisholm 1911; Anon 2017). A colour photograph of the specimen (sent by P. Campbell, Natural History Museum, London) was made available to the first author, and the ‘5N’ dorsal pattern was confirmed. This appears to represent an isolated population of D. scabra. Specimens of D. scabra (NMW 9964/6, ‘5N’) and D. confusa (NMW 9964/7, ‘5L’) from ‘Sudan’, donated to the Naturhistorisches Museum Wien in 1884 by ‘Emin Bey’, may have been collected nearby in South Sudan’s East Equatoria state, or possibly in the vicinity of Wadelai. Emin Pasha was governor of Equatoria (a region in southern South Sudan that included a portion of north-western Uganda), then a province of Anglo-Egyptian Sudan; he retreated to Wadelai in 1885 in the face of the Madhist advance (Chisholm 1911; Anon 2017).
ZFMK 32479 from 50 km north of Kutum, North Darfur State, western Sudan is a faded male specimen with a damaged head, referable to D. scabra on account of the following characters: ‘5N’ dorsal pattern with dark oval saddles and whitish interspaces, pattern cycles 50, SVL 447 mm, tail length 81 mm, SVL/tail ratio 5.5, ventrals 227, subcaudals 66, midbody scale rows 23. This locality is outside the study area and the only other known record of D. scabra in the north of Africa. It is not associated with the Nile River and represents an extremely isolated population, possibly a relic of a former widespread distribution (see discussion below under D. bazi). The nearest conspecific (RMNH 24399) is from near Kosti about 880 km ESE.
Dasypeltis bazi Saleh & Sarhan
Egyptian Egg-eater
Figs 7A–B, 8, 9A–B, 10–12
Rachiodon abyssinicus lapsus for abyssinus (not Duméril, Bibron & Duméril) Lichtenstein & Von Martens, 1856: 28 (“Oberaegypten” = Upper Egypt).
Rachiodon scaber (not Linnaeus) Jan, 1863: 106 (Cairo); Panceri, 1878: 22 (Egypt).
Dasypeltis scaber (not Linnaeus) Gasco, 1876: 119 (Middle Egypt).
Dasypeltis scabra (not Linnaeus) Anderson, 1896: 114, 1898: 278 (Fayum); Gans, 1959: 141 (part, Egypt, Fayum, Upper Egypt), Pl. IV, Figs 5& 6, Pl. VI, Fig. 1; Gans, 1961: 74 (part), Pl. 1, Figs A & B (Egypt); Marx, 1968: 33 (Egypt); Rehak & Osborn, 1988: 275 (Faiyum); Saleh, 1997: 146 (Faiyum); Baha El Din, 2006: 232 (Fayoum); Geniez, 2015: 210 (Fayoum).
Dasypeltis cf. scabra Sindaco et al. 2013: 93 (part, Egypt).
Dasypeltis “scabra” Göthel, 2015a: 19 (part) & 2015h: 86 (part).
Dasypeltis bazi Saleh & Sarhan, 2016: 31. Type locality: Abu-Gandir, Faiyum Governorate, Egypt, holotype AUZC-R09458.
Diagnosis. Assigned to the genus Dasypeltis on account of its slender form, possession of (usually 3–4) rows of reduced, oblique, keeled and serrated lateral scales (little or no serration in D. inornata) and head barely distinct from the neck (moderately distinct in D. fasciata). Distinguished from other congeners by its colour pattern consisting of: large cranio-caudally elongate dark brown saddles, usually bifurcated at each end with a fairly shallow but distinct notch, the saddles about 1.5 to 2 times as long as the pale interspaces between them (similar to D. loveridgei comb. nov., but this species has mostly transversely enlarged and distinctly hourglass-shaped saddles which are mostly about equal in length to the pale interspaces; similar but less elongate saddles in some populations of D. scabra, e.g. Fig. 5G & H); low numbers of pattern cycles 38–49 (lower than all other [patterned] species/populations except D. taylori sp. nov. and a few specimens of D. scabra; D. inornatus always uniform brown; D. gansi uniform or weakly marked); flanks decorated with dark brown spots or blotches (similar in D. loveridgei comb. nov.; and some D. scabra, e.g. 5G & H) rather than vertical bars as in most other (patterned) Dasypeltis species/populations; and venter immaculate white to cream-white apart from a few dark markings at the edges in some specimens (extensive dark markings in D. abyssina and many D. scabra, often extensive grey stippling in D. medici); together with a combination of the following characters: Ventrals 213–229 in males (mostly higher than D. taylori sp. nov. 196–216, lower than D. arabica sp. nov. 236–244), 215–232 in females (lower than D. abyssina 241–271, D. arabica sp. nov. 239–254, D. gansi 235–259, D. latericia 234–262; mostly lower than D. crucifera sp. nov. 231–247); Subcaudals 59–61 in males (lower than D. inornata 81–92, D. fasciata 71–90, D. medici 69–109, D. gansi 68–83, D. palmarum 68–77, D. parascabra 67–74, D. latericia 66–86, D. arabica sp. nov. 63–65), 51–63, 70 in females (mostly lower than D. inornata 69–84, D. fasciata 64–84, D. palmarum 62–77, D. medici 61–90); Tail of moderate length: SVL/tail length 5.0–5.9 in males, 5.1–7.2 in females (in study area, tail long in: D. medici 3.5–4.7 males, 4.5–5.7 females; D. fasciata 4.7–5.2 males, 4.8–6.4 females; see also Gans 1959); Postoculars 2 on either side of head (often one, especially in western populations, of D. atra); Nasal divided below the nostril (undivided in D. sahelensis and D. parascabra); Supralabials 7 (3rd and 4th enter orbit) on either side of head (usually 6[2,3] in D. loveridgei comb. nov.). Half the specimens examined had longitudinally divided anal plates; this plate is usually undivided in other species in the genus.
Description of BM 97.10.28.575 from Fayoum. (at Natural History Museum, London). Adult female (stretched oviducts and three eggs observed) with a total length of 692 mm, snout-vent length 602 mm, tail length 90 mm (Gans 1959) (ratio SVL: tail length 6.7, ratio total length: tail length 7.7) from Fayoum [about 29º18’30”N, 30º51’00”E], collected by A.R. Birdwood (Gans 1959, Pl. VI, Fig. 1; Figs 7 & 8). This is the same specimen discussed and illustrated by Anderson (1898, Pl. 34; see discussion under Remarks below) as D. scabra.
Head (somewhat dehydrated) small and barely distinct from the neck; snout rounded, rostral shield barely visible when viewed from above. The elongate nasal scale (about twice as long as high) is divided below the nostril (which is situated in the anterior half of the nasal); no loreal present. Three internasals between the rostral and paired prefrontals, but the larger, subcircular, central internasal appears to be an aberration. Each prefrontal is larger than an internasal (and the paired prefrontals together are much larger than all three internasals together), but they are of equal length (cranio-caudal orientation). Inter-prefrontal sulcus moderately marked. Frontal large, broadest anteriorly, 1.61 times as long as broad, smooth except for a single row of pits on the lateral borders, and of similar length to a supraocular. Frontal followed by a pair of large parietals, each one similar in size to the frontal. A single preocular and two postoculars present on either side of head. Temporal formula 3+3 (left), 2+3 (right). Seven supralabials on either side, the 3rd and 4th bordering the eye. First pair of infralabials in contact behind the mental, followed by a pair of large and elongate scales on the throat, and another pair of much smaller scales in contact with the first ventral plate. Eyes moderate in size, their greatest diameter subequal to the distance from tip of snout to anterior part of eye socket.
Dorsal scales strongly keeled, unicarinate, in 24 rows at midbody, the third, fourth and fifth rows laterally (counted from the first row in contact with the ventral plates) are denticulate (serrated), reduced in size and obliquely positioned; vertebral scales not enlarged. There are 227 smooth ventrals, anal plate divided, with 55 pairs of smooth subcaudals. After preservation in ethanol the dorsal colouration is pale brown, with 42 large, cranio-caudally elongate, medium-brown saddles (with darker edges, especially on their anterior and posterior ends) from nape to base of tail (i.e. above vent), most of which are about 1.5 to 2 times longer than wide, and bifurcated at each end with a fairly shallow but distinct notch. Inter-saddle region cream-white, distinctly paler than the rest of the dorsal surface. Lateral markings are in the form of medium-sized, medium-brown spots or blotches, with darker edges, on either side between the saddles. Dark vertebral blotches continue along the tail, but lateral markings here are narrow and elongate. There are extensive dark (brown) markings on the top and sides of the head (including the labials), including a pair of elongated stripes on either side of the frontal, a pair of anteriorly-joined teardrop-shaped markings (blunt anteriorly) on the front of the parietals, and a pair of larger, similar markings on the posterior part of the parietals which extend onto the neck. The saddle on the nape has a spike-like anterior extension which terminates bluntly on the medial-posterior part of the parietals where it is in contact with the larger pair of (posterior) parietal markings. The latter are also narrowly fused with the smaller pair of parietal markings. Venter immaculate cream, apart from a few small brown spots and flecks extending from the outer edges of the ventrals to the outer scale rows.
INSERT FIGS 7 & 8
Description of additional Egyptian material. There are six additional specimens (one is a skeleton which we assign to this species) of D. bazi from Egypt, mostly with inexact localities, but potentially all collected at or near Fayoum (see discussion below).
ZMB 1861 (at Museum für Naturkunde, Berlin), adult male (left side inverted hemipenis examined), from “Oberaegypten” (= Upper Egypt, a strip of land on either side of the Nile River from about Memphis just above the Nile Delta southwards to Lake Nasser in Nubia, which may include parts of northern Sudan) collected by Karl Richard Lepsius (born 1810, died 10 July 1844), a Prussian Egyptologist who was leader of an expedition to Egypt conducted between 1842 and 1846 (Figs 9–11). This expedition visited places from Memphis in the north traveling along the White and Blue Nile rivers into central Sudan. The specimen, in remarkably good condition, appears to be the same snake referred to by Lichtenstein & Von Martens (1856).
ZMB 51552, adult female (left side of tail base dissected open, no hemipenis) from “Oberaegypten” (= Upper Egypt), collected by the French physician Dr Alfred Peney (1817–1861), who was together with K.R. Lepsius in Egypt and Sudan; this specimen was apparently later donated to Albert von Barnim and Robert Hartmann who visited Peney on an expedition to Egypt, Nubia and eastern Sudan between 1860 and 1861. Barnim and Hartmann are mentioned on the jar label as donors for this specimen to the “Anatomisch-Zoologisches Museum” in Berlin, and the specimen was incorporated into the Zoologisches Museum Berlin in 1888.
ZMB 5865, adult male (inverted hemipenes present, F. Tillack in litt.) from “Aegypten” (= Egypt), purchased from Mr “Keitel”; specimen catalogued around 1867 by W.C.H. Peters as “Rachiodon abyssinicus Dum. Bib.”.
NMW 26879/1, adult female (no spines in tail, tail distinctly tapered at base) from “Omgebung v. Cairo” (= surroundings of Cairo), donated by Franz Steindachner in January 1874.
NMW 962, mounted adult skeleton with skull (few teeth present), from “Kairo” (= Cairo), donated or collected by Franz Steindachner in 1880, labelled “Dasypeltis scabra”.
MNHN 1847, adult male (left side hemipenis everted) from “Egypt”, collected by Baron Alphonse Marie Delort Gléon Leopard (1843–1899).
Size: ZMB 1861, male, 455 mm SVL + 79 mm tail length = 534 mm total length; ratio SVL: tail length 5.8, ratio total length: tail length 6.8; ZMB 5865, male, 427 + 78 = 505 mm, ratios 5.5 and 6.5; MNHN 1847, male, 410 + 70 = 480 mm, ratios 5.9 and 6.9; ZMB 51552, female, 503 + 70 = 573 mm, ratios 7.2 and 8.2; NMW 26879/1, female, 461 + 61 = 522 mm, ratios 7.6 and 8.6.
Scalation: All have similar scalation and general appearance to BM 97.10.28.575, except: Nostril on right side of NMW 26879/1 situated in about middle of nasal scale (not in anterior part thereof). All have a pair of internasals (not three) between rostral and paired prefrontals, and these are (individually and in combination) smaller than the prefrontals (smaller but of equal length in ZM 51552). Inter-prefrontal sulcus weakly marked in ZMB 5865 and ZMB 51552. Ratio frontal length: frontal width: 1.54 (ZMB 1861), 1.44 (ZMB 5865) and 1.40 (MNHN 1847) in males; 1.38 (ZMB 51552) and 1.43 (NMW 26879/1) in females; frontal smooth in ZMB 51552, covered by small pimple-like protrusions in MNHN 1847. Frontal slightly shorter than each supraocular in NMW 26879/1. In ZMB 1861 the pair of parietals is fused medially, with only the ends separated. Temporals 2+3 on both sides. In NMW 26879/1 and ZMB 1861 the pupils are clearly vertically elliptical. Midbody scale rows 23 or 24 (NMW 26879/1, ZMB 51552). Reduced, oblique, serrated lateral scale rows: 3–5 (MNHN 1847, ZMB 5865) and 3–6 (NMW 26879/1, ZMB 1861, ZMB 51552), but all except NMW 26879/1 and ZMB 51552 also have the scales of the second row weakly serrated although not reduced in size like the other serrated rows, nor obliquely oriented. Ventrals in males 219 (ZMB 5865), 229 (ZMB 1861, MNHN 1847); in females 228 (ZMB 51552), 234 (NMW 26879/1). Anal plate undivided in MNHN 1847, ZMB 1861 and ZMB 51552. Subcaudals in males 59 (MNHN 1847, ZMB 1861), 60 (ZMB 5865); in females 50 (NMW 26879/1), 51 (ZMB 51552).
Colour pattern: Basal colour dorsally in MNHN 1847 is grey, not pale brown. Saddles on back 39 (ZMB 5865), 48 (ZMB 1861, ZMB 51552), 49 (MNHN 1847); about 48 in NMW 26879/1 which is largely faded. Along the tail the lateral markings are in the form of small dark spots/blotches in MNHN 1847, NMW 26879/1 and ZMB 1861. Brown forward-pointing V-shaped markings on back of head and nape in MNHN 1847; distinct, backwards-curving, narrow pale band at back of head, bisected by spike-like extension of a dark and mostly blunt V-shaped marking (almost band-like) on nape in ZMB 5865; ZMB 1861 and ZMB 51552 similar to previous specimen but pale band not interrupted, and the latter with a longer and sharper anterior extension of the nape marking.
INSERT FIGS 9–11
Description of ZFMK 38415 (Dasypeltis cf. bazi). Adult male (600 mm total length) from Erkowit, north-eastern Sudan [18º46’N, 37º07’E], collected dead-on-road (head damaged) in Spring 1982 by Gerhard Nikolaus (Fig. 12).
Ventrals 229, subcaudals 67, midbody scale rows 25, supralabials 7 (3rd and 4th enter orbit) on either side of head, one preocular on either side, two postoculars on either side, 3+4 temporals on either side, 48 pattern cycles. Photographs (e.g. Fig. 12) of the specimen show a dorsal pattern similar to that of BM 97.10.28.575; venter also similar, but yellowish-cream; dorsal and ventral scales, including their arrangement on the throat, similar to holotype; reduced, oblique, serrated lateral scale rows 3–6, but second row (scales similar in size to central dorsals) weakly serrated although not reduced or oblique; frontal similar in shape to that of the holotype, smooth except for 1–2 rows of pits laterally, of similar length to a supraocular, but slightly larger than each of the parietals.
INSERT FIG. 12
Size. Largest male ZMB 1861 (“Upper Egypt”) 455 + 79 = 534 mm [The largest specimen identified as a male by Saleh & Sarhan 2016 measured 482 + 75 = 557 mm, but its sex requires confirmation—see below]. Largest female BM 97.10.28.575 (Fayoum, Egypt) 602 + 90 = 692 mm.
Distribution. Dasypeltis bazi appears to be restricted to an area of about 300 km2 along the El Buts channel (near the village of Ibshwai) in the Fayoum Depression, which is situated about 100 km south-west of Cairo and 40 km west of the Nile River, at elevations of 32–52 m below sea level (Saleh & Sarhan 2016). Apart from sites in the Fayoum, there are a records of this species from only one other specific locality in Egypt, namely Cairo (see discussion below), but its occurrence there was not confirmed by Saleh & Sarhan (2016). Cairo is now a densely populated area, and if these records are accurate, the species may have been extirpated there. Although it has been suggested that it occurs elsewhere in Egypt along the Nile River (e.g. Anderson 1898; Baha El Din 2006), this was also not confirmed by Saleh & Sarhan (2016) who investigated potential habitats in various parts of the country.
Historical records. Based on the literature, D. bazi is a poorly known species. The first record of an egg-eating snake from Egypt is contained in Lichtenstein & Von Martens’ (1856) list of the reptiles and amphibians in the collection of the Royal University of Berlin. He recorded a specimen of Rachiodon abyssinicus (lapsus for abyssinus) from “Oberaegypten” (= Upper Egypt). Jan (1863) later recorded Rachiodon scaber (= D. scabra) from Cairo with reference to a specimen in the Museo Civico di Storia Naturale di Milano. However, the latter specimen was not illustrated, nor were any morphological data provided, and it is no longer to be found at the museum in Milan (S. Scali in litt. 26 September 2014). No other mention has ever been made of the presence of this species in Cairo, a densely populated area, so it may have been extirpated there. In an account of his travels in Egypt, Gasco (1876) recorded eight specimens of Dasypeltis scaber (= D. scabra) from Middle Egypt (= Nile Valley between the Nile Delta and Asyut or Qena Bend), collected in 1873–1874. The title of Gasco’s (1876) paper, the second of two reports on his travels in Egypt, actually refers to the oasis of Fayoum and excursions in the vicinity of Cairo, although it also alludes to hunting on Lake Menzala on the Nile Delta. Anderson (1896: 114) noted that “Count Peracca has been so good as to ascertain from Prof. Costa that two specimens brought back from Egypt by Prof. P. Panceri, the companion of Gasco, one a skeleton and the other in alcohol, are preserved in the Naples Museum. Panceri (1878) did in fact refer to two such specimens (number 3219, alcohol specimen; 3520, skeleton and teeth), as Rachiodon scaber, in his published catalogue of the collection in the Comparative Anatomy section of Università degli Studi di Napoli Federico II. It is not known whether the two specimens from Cairo in the Vienna collection, as mentioned above, were a part of the series alluded to by Gasco and Panceri, but the dates involved do coincide.
Subsequently, in Zoology of Egypt, Anderson (1898: 279) wrote: “After much correspondence with those in Egypt who have been assisting me in my researches, at last, through the efforts of Mr. A. R. Birdwood, after I had almost given up all hope of finding this species, I received the specimen herein recorded alive and am thus enabled to give a figure of it from life.” According to Anderson (1898: 279) the specimen was collected in “the Fayum” (variously spelled as “Faiyum”, “Fayoum”, “Al Fayyum” and “El Faiyūm”), which probably refers to the oasis in general rather than the city of Fayoum itself; the area is also referred to as the Fayoum Depression. Anderson (1898, table) was of the opinion that D. scabra also occurred in the “Valley of White Nile”. In fact, Anderson’s (1898: 279) comment about the “wide distribution” of ‘D. scabra’ and his suggestion that the species occurs “northwards along the valley of the Nile to within about 140 miles [= 225 km] of the Mediterranean [Sea]” is somewhat misleading because apart from the ‘Nil Blanc’ record (a syntype of R. abyssinus; see below) of D. scabra far to the south, there does not appear to have been (nor is there now) any other specific documented evidence to suggest that the Fayoum population was in fact contiguous with other Dasypeltis populations. For his Fayoum specimen Anderson (1898) provided scalation and size data, line drawings of the head and anterior part of the body, and an illustration (painting in colour) of the snake in the process of swallowing a pigeon’s egg.
Anderson’s specimen (BM 97.10.28.575) was later examined by Gans (1959) who remarked on the uniqueness of its dorsal pattern. Gans (1959) also examined three more specimens (also listed and examined here) from Egypt and “Upper Egypt” (ZMB 1861, this is probably the snake mentioned by Lichtenstein & Von Martens 1856). One additional specimen (ZMB 51552) from “Upper Egypt” recorded here can be added to the list of old museum specimens. These non-specific localities could conceivably refer to specimens collected at other localities apart from the Fayoum Depression, perhaps along the Nile River itself or possibly in one of the country’s other oases.
Rehak & Osborn (1988: 275) later recorded a specimen “collected by natives in Faiyum, September 1982” and noted that it “seems extremely rare in Egypt”. Saleh (1997) subsequently provided a photograph of a specimen he collected in Fayoum, again illustrating the much-elongated saddles with spots/blotches on the flanks. Although Saleh (1997) claimed that his specimen from Fayoum was the first to be collected in almost a century, Rehak & Osborn (1988) had in fact documented a specimen collected there about 90 years after Anderson’s (1898) report. Baha El Din (2006) was of the opinion that the only certain locality for D. scabra in Egypt is the Fayoum Depression, but it is also likely to occur in parts of the Nile Valley. He added that Gasco’s (1876) locality of “Middle Egypt” may be in reference to Fayoum, which is located within the northern parts of this region. Baha El Din (2006) also noted that animal collectors from Abu Rawash (30o01’55”N, 31o04’30”E; near the southern edge of the Nile Delta, about 80 km north-east of Fayoum) claimed to find specimens at pigeon breeding houses. If it does in fact occur in the vicinity of Abu Rawash, this would be the most northerly known locality for the species and the most north-easterly extent of the range of the genus Dasypeltis.
The new species was therefore, prior to 2016, known to science from only 16 specimens recorded over a period of about 170 years (first specimen collected by Karl Lepsius in about 1844, see above). Saleh & Sarhan (2016) reported in detail on eight specimens collected at Fayoum in the period 1995–2015 (presumably including Saleh’s [1997] specimen) and also mentioned another 17 specimens collected there since 1995 but released. There are no other records of egg-eaters from Egypt. Venchi & Sindaco’s (2006) checklist of the reptiles of the Mediterranean region did not add any additional records of Dasypeltis. Watkins-Colwell et al. (2010) reported on reptiles recorded during three Yale University expeditions to Egypt in the first half of the 1960s, which included several sites along the Nile River and the Fayoum Oasis, but no Dasypeltis were recorded. Ibrahim (2013) recently reported on the herpetofauna of the Suez Canal Zone, surveyed between 1999 and 2012. Despite the fact that habitat changes in the area at and near the canal (which includes associated freshwater irrigation canals) have allowed ‘invasive’ species to expand their ranges, and that an extensive web of irrigation canals has provided passage for several species to move from the Nile River to the Suez Canal, no specimens of Dasypeltis were recorded. If Dasypeltis occurred in the Nile Delta itself, for which there is no actual evidence, this too would allow easy access to the Suez Canal.
Natural history. Saleh (1997: 146) collected a specimen from Fayoum “in an area with dense wild vegetation, not far from houses at the edge of cultivated land”. Saleh & Sarhan (2016) later provided a detailed description of the habitat of the Fayoum population. All specimens were collected along the El Buts channel, a relict section of the Fayoum ancient natural drainage system. This habitat comprises a fairly deep gorge in the thick and silty Nilotic deposits in the central region of Fayoum Depression. Terraces occupy the sides where small plots of crops are grown (Saleh & Sarhan 2016). Anderson (1898: 280) noted: “On Plate XXXIX. it [his Fayoum snake—perhaps depicted while in captivity] is represented in the act of swallowing a pigeon’s egg, but fowls’ eggs are even within its capacity.” Saleh & Sarhan (2016) later determined that several bird species that produce suitably small eggs occur in sympatry with these snakes at Fayoum. In addition, they noted that traditional pigeon towers and chicken hatcheries also provide a source of eggs. Based on the presence of three large eggs observed in the abdomen of BM 97.10.28.575, this species is, like other congeners for which data is available (e.g. Broadley 1990; Spawls et al. 2002), oviparous.
Mimicry. Dasypeltis bazi has a dorsal colour pattern similar to the saw-scaled viper Echis pyramidum Geoffroy Saint-Hilaire (Schleish et al. 1996; see photographs in Dobiey & Vogel 2007) which it appears to mimic. Like Echis Merrem, it has similar serrated lateral dorsals which it may engage for defensive display by coiling and rubbing together to produce a rasping sound (scale stridulation, Schleish et al. 1996). The type series of Echis pyramidum lucidus Cherlin, 1990 was collected in the Fayoum Oasis (Cherlin 1990). A specimen of the latter subspecies from Biskra in Algeria (www.megasphera.cz/africanvenomoussnakes/, accessed 11 June 2014) has a dorsal pattern almost identical to D. bazi (e.g. Fig. 7A-B). Echis pyramidum was also collected in the Fayoum Oasis during the Yale University Expedition (Watkins-Colwell et al. 2010). It is one of several species of snakes that occur in the same habitat as D. bazi (see Saleh & Sarhan 2016). Anderson (1898) suggested that the similarity of the Fayoum snake to Echis carinatus (Schneider) (= ? E. pyramidum) might explain why this egg-eater is so seldom observed or recognised, but Saleh (1997) noted that the Fayoum Dasypeltis that he obtained was not particularly similar to this viper. Gans (1961, Pl. 1) illustrated the similarity between these two species using ZMB 5865 (‘Egypt’) and a specimen of ‘E. c. carinatus’ (CAS 38722) from Cairo.
Biogeography. Dasypeltis bazi appears to have a relict distribution, the result of a once extensive African savannah being reduced and fragmented following dessication of the region during the Holocene (Kuper & Kröpelin 2006). This is discussed in more detail below (see Discussion).
Conservation. The Fayoum Oasis is a depression or basin linked to the Nile River by a 24-km-long canal called the Bahr Yussef. In prehistory the oasis contained the large freshwater Lake Moeris, which received water when the Nile was in high flood, but it is now a much smaller saltwater lake referred to as Birket Qaran (= Qaroun Lake). In 2300 B.C.E. the natural waterway from the Nile to the lake was widened and deepened to control flooding, regulate water levels during the dry season, and provide the area with irrigation for crops. Unlike other oases in Egypt, which have spring water sources, cultivated land in the Fayoum is formed by fertile mud from the Nile transported there via the Bahr Yussef. The scarcity of Dasypeltis records from the Fayoum is perhaps not too surprising considering the high levels of human activity (e.g. crops and housing) in the region. Saleh & Sarhan (2016: 37) noted: “Through centuries of human occupation and intensive agriculture, the Faiyum natural drainage system was fundamentally modified and almost totally obliterated.” Proposed tourism developments will impose further threats to natural habitat in the area (Khalifa & El-Khateeb 2011).
Despite the benefits provided to the human population of Egypt by the building of the Aswan High Dam, which created Lake Nasser, this dam has permanently altered the ecology of the Nile River region, with some areas now under water and others changed from desert into cultivated fields or wetlands (Watkins-Colwell et al. 2010). Although the establishment of vegetated areas has resulted in some species extending their ranges, e.g. inland from the Nile Delta (Baha El Din 2006; Watkins-Colwell et al. 2010), this does not appear to have benefited Dasypeltis. Since the construction of the dam and the resultant low flow rates, the Nile Delta no longer floods and has been converted into an intensive agricultural area (Van As et al. 2012). In the past specimens may have been introduced downstream—even from as far away as Sudan—during floods, on rafts of floating vegetation, but this no longer occurs. Although the delta is still an important area for a number of animal species, including large numbers of migratory birds (Mockrin & Thieme undated), no egg-eating snakes have been recorded there.
Considering the scarcity of records as indicated above, D. bazi may be a naturally rare species. Anderson (1898: 280) noted: “It seems remarkable that a snake of this kind, so specialized as regards its food-supply, should have been unknown to the ancient Egyptians. There is no evidence, so far as I have been able to discover, that they were acquainted with it, unless the figure, said to occur on the monuments, of a hawk-headed serpent with an egg in its mouth may have been derived from some knowledge of it.” The Egyptian god Cneph is in fact occasionally represented as a serpent with an egg in its mouth (Deane 1833). Anderson (1898) also noted that he had made a special effort to obtain specimens, yet he only eventually secured a single snake, from Fayoum. Rehak & Osborn (1988) also recorded a single specimen and considered Dasypeltis as extremely rare in Egypt. This specimen was apparently collected by a Fayoum villager in September 1982 but was purchased from an animal dealer in Cairo (see Saleh & Sarhan 2016). Saleh (1997) and Baha El Din (2006) classified the Egyptian population of ‘D. scabra’ as regionally Critically Endangered. Baha El Din (2006: 234) noted: “The impact of the large-scale ecological changes that took place in Fayoum since the beginning of last century is unknown, but probably reduced available habitat. The species is threatened by over-collection by professional animal collectors [in Egypt], who target the species.” This suggestion about over-collecting appears to be based on his comment about animal collectors from Abu Rawash. Saleh & Sarhan (2016: 43) concluded that D. bazi was “saved from certain extinction by the development [since the mid-Holocene] of artificial breeding of pigeons and chickens in Faiyum” which provided a source of eggs.
The Erkowit specimen may represent an isolated relict population of D. bazi in north-eastern Sudan. This population may in the past have been part of a more widespread population of D. scabra that became isolated following aridification of the Sahara, resulting in the evolution of a new species, D. bazi. The Egyptian and Erkowit populations themselves may then have become isolated, accounting for their large current-day separation. Habitat at Erkowit has been negatively affected by the impacts of a human population that has grown considerably in the last 60-odd years, with associated removal of wood, pressure from livestock grazing and browsing, and a warming climate (Vetaas et al. 2012). This environment has suffered continuous change from semi-evergreen woodland to open shrubland similar to that of the surrounding desert, and the area can no longer be regarded as a ‘mist oasis’ dominated by Afromontane and Mediterranean plant species (Vetaas et al. 2012). Therefore, there is some uncertainty about the conservation status of the Erkowit population.
Based on a global evaluation of its conservation status using IUCN (2012, 2016) criteria—and considering the Fayoum Oasis population as the only surviving population of this species (see above)—it is evident that D. bazi should be classified globally as Endangered [B1ab(i, ii, iii)] based on its small geographical range (known from an area of 300 km2 [Saleh & Sarhan 2016], extent of occurrence <5000 km2 [the entire oasis is about a third of this size]) which is severely fragmented, with an inferred continuing decline in its extent of occurrence, area of occupancy and quality of habitat (see Baha El Din 2006; Khalifa & El-Khateeb 2011; Saleh & Sarhan 2016).
Remarks. Although Anderson (1898) did not identify his Fayoum snake by a museum accession number, he did mention that reptile and amphibian specimens collected and discussed by him were lodged in the British Museum (now in the Natural History Museum, London), the Museum of Egyptian Government School of Medicine (Cairo) and a few museums on the European mainland. Baha El Din (2006: 233) actually identified Anderson’s specimen as “BMNH 97.10.28.575”. Based on the illustrations, measurements and scalation details provided by Anderson for his Fayoum snake, it is apparent (see below) that his specimen is indeed the same snake preserved at the Natural History Museum in London and labelled as BM 97.10.28.575, which was examined by both Gans (1959, see Pl. 6, Fig. 1) and the first author. Firstly, the body shape, and shape and proportion of dorsal markings in Anderson’s (1898) Pl. 34, Fig. 3 are remarkably similar to the specimen examined (Figs 7 & 8), including the first saddle on the nape with its narrow finger-like forward projection. Secondly, measurements of SVL and tail length by Anderson (602 + 95 mm) and Gans (602 + 90) are similar. Thirdly, there are general similarities with regard to scalation. Anderson’s ventral count of 230 is similar to the count of 227 determined by both Gans and the first author. The slight difference would be accounted for if Anderson included in his count the three pairs of scales between mental and first ventral. Some of Anderson’s scale counts are in exact agreement with those obtained by Gans and this author: 24 midbody scale rows, one preocular and two postoculars. Anderson’s subcaudal count of 49 is slightly lower than the 54 counted by Gans and the 55 counted for the present study. However, because subcaudals in Dasypeltis are paired—and these paired scales are not always clearly aligned—it is possible that Anderson under-counted, thus minimising this count. Also, while Anderson recorded eight supralabials with 4th and 5th entering the orbit, both Gans and the first author recorded only seven supralabials with 3rd and 4th entering the orbit. However, an examination of Anderson’s (1898) Pl. 34, Fig. 3, representing a lateral view of the right side of the head and neck, clearly shows seven supralabials with the 3rd and 4th entering the orbit. It seems that Anderson may have inadvertently or erroneously included the rostral in his count, which would have created the impression that the ‘4th’ and ‘5th’ supralabials entered the orbit. Anderson provided a temporal formula of 3+3, while Gans recorded 2+3, but the first author observed 3+3 on the left side of the head and 2+3 on the right.
A few inaccuracies were noted in Saleh & Sarhan’s (2016) diagnosis of D. bazi. Firstly, these authors state that in D. bazi the frontal shield is longer than wide, but as long as wide in other species of Dasypeltis. In fact the frontal is longer than wide in most if not all congeners (e.g. Gans 1959; Broadley 1990; Trape & Mané 2006; Trape et al. 2012; figures in this paper). Also, the temporal arrangement of 2+3 (two anterior and three posterior temporals on either side of the head) is common in the genus and the norm for several species (Gans 1959; Broadley 1990; this paper); while one preocular on either side of the head is the norm in all species, even though there are occasionally two or three such scales in some species (Gans 1959; this paper). Elsewhere in their paper Saleh & Sarhan (2016) recorded low numbers of longitudinal scale rows at midbody (20–21, one specimen with 22). Egyptian specimens of D. bazi examined for the present study had counts of 23–24. Saleh & Sarhan (2016) also counted ‘anterior’ and ‘posterior’ longitudinal scale rows, and their ‘anterior’ scale row counts (presumably counted halfway between tip of snout and midbody) of 21–25 were closer to the values obtained in the present study. It is possible that they did not use the same method for counting these scale rows as other authors, especially that proposed by Gans (1959) and used in this study, and therefore their comparisons with other species are not feasible. Saleh & Sarhan (2016) added to the confusion when comparing ‘dorsal scale rows’ between species by using the total range of anterior, midbody and posterior scale row counts for D. bazi (i.e. ‘18–24’; but 18–25 in their Table III) and comparing this with what is almost certainly only midbody scale rows recorded by other authors for other species. Lastly, a specimen (AUZC-R12006) identified as a male by Saleh & Sarhan (2016) has a very high ventral count of 250 (and low subcaudal count of 55), and may be a female; the next highest ventral count for a male D. bazi is 229 (see above).
Dasypeltis taylori Bates & Broadley sp. nov.
Taylor’s Egg-eater
Figs 13–14
Dasypeltis scabra var. Medici (not Bianconi) Mocquard, 1888: 131 (‘Comalis’ = Djibouti).
Dasypeltis scabra (not Linnaeus) ? Scortecci, 1930: 213 (Assab, Eritrea) (material not examined); Gans, 1959: 141 (part), Pl. VI, Fig. 6 & Pl. VII, Fig. 1; Balletto, 1968: 268 (part); Lanza, 1983: 223 (part), 1990: 436 (part); Ineich, 1999: 35 (Djibouti).
Dasypeltis scaber (not Linnaeus) Parker, 1949: 66 (Haud, Ogo and Hargeisa, Somalia).
Dasypeltis “scabra” Göthel, 2015a: 19 (part) & 2015h: 86 (part).
Holotype. BM 49.2.2.2 (at Natural History Museum, London), a subadult male (Gans 1959) from Haud [former British Somaliland, 08°20’N, 46°00’E at 2100 ft = 640 m] in northern Somalia, collected by Colonel R.H.R. Taylor on 20 April 1932 (Fig. 13).
Paratypes. Seven specimens: SOMALIA (Somaliland). Allotype. BM 49.2.2.3 (at Natural History Museum, London), a subadult female (Gans 1959) with the same data as the holotype, but collected on 1 May 1932 (Gans 1959, Pl. VI, Fig. 6); BM 49.2.2.1 (subadult male) with the same data as the holotype; BM 49.2.2.4, adult female from Hargeisa District [9°35’N, 44°05’E at 4000 ft = 1220 m a.s.l.] collected 29 May 1932; BM 1992.501, male from 2.5 km WSW of Hargeisa [09°34’N, 44°03’E at 1275 m a.s.l.] collected 29 March 1992 by M.R.K. Lambert; BM 49.2.2.5, adult male from Ogo [10°10’N, 42°50’E at 4500 ft = 1370 m a.s.l.] collected 6 August 1933; BM 51.1.7.69, skin of a male from Garadak [09°29’N, 46°18’E at about 1100 m a.s.l.]. DJIBOUTI. MNHN 84–707, adult female from ‘Comalis’ [= Djibouti] (Fig. 14).
Diagnosis. Assigned to the genus Dasypeltis on account of its slender form, possession of (usually 3–4) rows of reduced, oblique, keeled and serrated lateral scales (little or no serration in D. inornata) and head barely distinct from the neck (moderately distinct in D. fasciata). Distinguished from other congeners by a combination of the following characters: Colour pattern usually consists of: large, usually closely-spaced, square to slightly rectangular saddles, separated by narrow, usually sharply defined, white crossbars (a variety of ‘5N’, Gans 1959, Pl. VI, Fig. 6); low numbers of pattern cycles 44–59 (≥62 in patterned species/populations of D. palmarum, D. fasciata, D. abyssina, D. atra, D. arabica sp. nov., D. inornatus always uniform brown; D. gansi uniform or weakly marked); flanks decorated with dark vertical bars (spots or blotches in D. bazi, D. loveridgei comb. nov. and some populations of D. scabra); and venter immaculate apart from a few dark markings at the edges (extensive dark markings in D. abyssina and many D. scabra); Frontal shield extensively pitted, States 3 & 4 (usually marginally pitted in most other species except D. medici, and some populations of D. scabra [especially in the south of its range] and D. loveridgei comb. nov., which are similar); Inter-prefrontal sulcus usually moderately depressed (usually weakly marked in most other species, but moderate to well marked in D. arabica sp. nov., weakly to moderately marked in D. bazi, and variable in D. scabra [often well marked in the south of its range]); Low numbers of ventrals: 196–216 in males (≥219 in: D. arabica sp. nov. 236–244, D. abyssina 226-247, D. crucifera 226, D. gansi 221–240, D. latericia 219–243, mostly lower than D. medici 215–254 and D. bazi 213–229), 208–222 in females (lower than D. abyssina 241–271, D. arabica sp. nov. 239–254, D. gansi 235–259, D. latericia 234–262, D. crucifera sp. nov. 231–247, D. fasciata 225–260, D. parascabra 224–237; mostly lower than D. palmarum 220–248 and D. bazi 215, 219, 224–232); Low numbers of subcaudals: 55–68 in males (lower than D. inornata 81–92, D. fasciata 71–90, D. medici 69–109; mostly lower than D. gansi 68–83, D. palmarum 68–77, D. parascabra 67–74, D. latericia 66–86), 44–52 in females (lower than D. inornata 69–84, D. fasciata 64–84, D. parascabra 57–64, D. palmarum 62–77, D. medici 61–90, D. latericia 59–72, D. gansi 59–73, D. arabica sp. nov. 53–61); Tail of moderate length: SVL/tail length 4.9–5.7 in males, 7.0–8.2 in females (in study area mostly greater than: D. medici 3.5–4.7 males, 4.5–5.7 females; D. fasciata 4.7–5.2 males, 4.8–6.4 females; see also Gans 1959); Nasal divided below the nostril (undivided in D. sahelensis and D. parascabra); Postoculars 2 on either side of head (often one, especially in western populations, of D. atra); Supralabials usually 7 (3rd and 4th enter orbit) on either side of head (usually 6[2,3] in D. loveridgei comb. nov.).
The new species is most similar to D. scabra, which usually has higher numbers of pattern cycles on the back—in surrounding countries/regions only two specimens, from southern Somalia (50–51 cycles) and a single snake from Sudan (54 cycles), have similarly low counts (Appendix 1).
Description of holotype The holotype (BM 49.2.2.2) is a subadult male with a total length of 202 mm, snout-vent length 172 mm, tail length 30 mm; ratio SVL: tail length 5.7, ratio total length: tail length 6.7.
Head small and barely distinct from the neck; snout rounded. Elongate nasal semi-divided (divided below the nostril), no loreal. Paired prefrontals much larger than paired internasals (individually and in combination). Inter-prefrontal sulcus moderately marked. Frontal large, slightly wider anteriorly, 1.51 times longer than broad, and fully pitted (State 4), slightly longer than each supraocular, and followed behind by a pair of large parietals each of which is larger than the frontal. There is a single preocular and two postoculars. Temporals 2+3. Seven supralabials, the 3rd and 4th bordering the eye. First pair of infralabials in contact behind the mental, followed by a pair of large and elongate sublinguals, and another pair of much smaller sublinguals in contact with the first ventral plate. Eyes moderate in size (with vertically elliptical pupils), their length subequal to the distance from tip of snout to anterior part of eye socket.
Dorsal scales strongly keeled, unicarinate, in 24 longitudinal rows at midbody, the third, fourth and fifth rows denticulate and (except for the 3rd row) obliquely placed. Vertebral scale row not enlarged. There are 198 smooth ventrals, the anal is entire and there are 60 pairs of smooth subcaudals. After preservation in ethanol the dorsal colouration is grey with 50 dark dorsal saddles between the nape and the base of the tail, separated by sharply defined white crossbars. Similarly dark, mostly elongate and wavy, lateral bars are linked to these crossbars on either side. Back of head with a distinct forward-pointing chevron; saddle on nape weakly forked posteriorly with a rectangular anterior prolongation. Venter immaculate cream-white apart from a few small brown spots on the outer edges of the ventrals.
INSERT FIG. 13
Variation in paratypes (including allotype, as indicated). The seven paratypes comprise four males and three females. The largest male (BM 49.2.2.5) measures 501 (408 + 83) mm, SVL/tail length ratio 4.9, total length/tail length ratio 5.9; one other male with measurements: BM 49.2.2.1 (188 + 34 mm, ratios 5.5 and 6.5). Largest female (MNHN 84–707) measures 545 (477 + 68 mm, ratios 7.0 and 8.0); other females: BM 49.2.2.4 (530 mm SVL), BM 49.2.2.3 (189 + 23 mm, ratios 8.2 and 9.2; allotype). The allotype and paratypes have similar scalation and general appearance to the holotype, except: Snout less rounded in BM 49.2.2.4. Inter-prefrontal sulcus weakly marked in BM 49.2.2.1 and 49.2.2.5. Frontal distinctly wider anteriorly in BM 49.2.2.4, BM 51.1.7.69, BM 1992.501 and MNHN 84–707; 1.25 to 1.53 times as long as wide; largely (not fully) pitted (State 3) in BM 51.1.7.69 and MNHN 84–707; and about as long as (not longer than) each supraocular in BM 49.2.2.5. Frontal about the same size as each of the parietals in BM 1992.501. One postocular (not two) on both sides of head in BM 49.2.2.4, and one on right side in BM 49.2.2.3 (allotype). Temporals 1+3 in BM 49.2.2.4 and 2+2 in BM 51.1.7.69. Six supralabials, the 2nd and 3rd bordering the eye, in BM 49.2.2.1. In BM 49.2.2.4 the pair of posterior sublinguals is largely separated by a more-or-less quadrangular supranumerary scale which is in contact with the first ventral plate. Length of eye almost equal to distance from tip of snout to anterior part of eye socket in BM 49.2.2.1 and BM 51.1.7.69, distinctly smaller than snout-eye distance in BM 49.2.2.4, 49.2.2.5 and MNHN 84–707. Midbody scale rows 23 in two specimens and 25 in two others (see Appendix 1); about 24 in BM 1992.501 (difficult to count) which has the third and fourth, and in some areas also the fifth, rows denticulate and oblique. Dorsal scales of the anal region distinctly denticulate in BM 49.2.2.4 and MNHN 84–707, feebly denticulate in BM 49.2.2.1, 49.2.2.3 (allotype) and 51.1.7.69. Ventrals 196-216 in males, 208–222 in females (see Appendix 1). Subcaudals 55–68 in males, 44–52 in females (tail truncated in BM 1992.501, male; and BM 49.2.2.4, female) (see Appendix 1). Dorsal pattern cycles 44–59 (see Appendix 1). In a flat skin (BM 51.1.7.69) the two halves of the vertebral saddles are not synchronised, forming an oblique pattern [Gans, 1959, Pl. VII, Fig. 1, erroneously given as ‘BM 51–1–7–59’]; and several dorsal saddles are oblique in BM 49.2.2.1. Dorsal colouration (excluding dark saddles and lateral bars) in preservative mostly rufous brown in BM 49.2.2.3 (allotype), a mixture of rufous brown and grey in BM 49.2.2.1 and 49.2.2.5, and pale brown in BM 49.2.2.4. Several lateral bars non-elongate and irregular in shape in BM 49.2.2.5. Pale crossbars between saddles in BM 49.2.2.4, 49.2.2.5, 51.1.7.69 and 1992.501 are thicker and less sharply defined than in the holotype [and allotype BM 49.2.2.3, as illustrated in Gans 1959, Pl. VI, Fig. 6]. Saddles are mostly rectangular and somewhat elongate in BM 49.2.2.5 and 1992.501, and in BM 49.2.2.4 they are rectangular at midbody but vary considerably in shape elsewhere, from slightly transversely enlarged and oblique, to oval and even ‘5I’-like (see Gans 1959). In BM 1992.501 several lateral bars are in contact with the saddles. Back of head with anterior part of chevron somewhat fragmented in BM 49.2.2.1 and MNHN 84–707, and bulbous in BM 1992.501; top of head somewhat faded in BM 49.2.2.3 (allotype); saddle on nape deeply forked posteriorly in BM 49.2.2.4 & 5, and always with a sharp anterior prolongation. Venter cream with large grey spots laterally, and smaller grey markings elsewhere, in BM 49.2.2.5; and whitish with 1–2 longitudinal rows of brown spots at the edges of the plates in BM 1992.501 and MNHN 84–707. No live specimens or photographic images of living snakes of this species are known to the authors.
INSERT FIG. 14
Etymology. The name of this new species is a patronym honouring Colonel R.H.R. Taylor, who collected about half of the type series while surveying the border between British Somaliland and Ethiopia in 1932–3.
Size. Largest male (BM 49.2.2.5 – Ogo, Somalia) 408 + 83 = 501 mm; largest female (MNHN 84–707 – Djibouti) 477 + 68 = 545 mm.
Habitat. Grasslands to Acacia–Commiphora bushland from 640 m to 1370 m (Figs 15 & 16). BM 1992.501 from 2.5 km WSW of Hargeisa was found on a limestone hillside (1275 m) with Vachellia etbaica Schweinfurth bushes.
INSERT FIGS 15 & 16
Distribution. Dasypeltis taylori sp. nov. has been recorded only from the arid interior of northern Somalia (Somaliland) (Figs 15 & 16) and in adjacent Djibouti (Fig. 3).
Mimicry. This species appears to mimic the saw-scaled viper Echis pyramidum Geoffroy Saint-Hilaire (see photographs in Dobiey & Vogel 2007 and Mazuch 2013). The racer Platyceps taylori (Parker), in the Borama area of Somaliland, has a dorsal pattern (see photograph in Mazuch 2013) remarkably similar to that of D. taylori sp. nov.
Biogeography. Dasypeltis taylori sp. nov. is separated from D. scabra populations in southern Somalia by the arid Ogaden plateau of eastern Ethiopia, an area from which no Dasypeltis have been recorded (e.g. Mazuch 2013). The highlands of east-central Ethiopia are occupied by D. atra, and those of north-western Ethiopia and southern Eritrea by D. abyssina. Dasypeltis taylori sp. nov. may therefore represent a relictual population of a once widespread D. scabra ancestor. Schätti & Ineich (2004) have discussed the radiation of the snake genus Platyceps Blyth, which is even more speciose than Dasypeltis in the Horn of Africa. Similar diversification occurs in the terrapin Pelomedusa subrufa species complex, where P. somalica Petzold et al. 2014 occurs in northern Somalia, P. gehafie (Rüppell) in Eritrea and eastern Sudan, and P. barbata Petzold et al. 2014 in Saudi Arabia and Yemen (Petzold et al. 2014). Typhlops calabressi Gans & Laurent has a very similar distribution to D. taylori sp. nov. (Broadley & Wallach 2009).
Conservation. In terms of the regional (study area) and global distribution of this species, using IUCN (2012, 2016) criteria, it appears to be widespread and should be regarded as Least Concern.
Remarks. Scortecci (1930) recorded “D. scabra” from Assab on the Red Sea coast of southern Eritrea; this specimen may be D. taylori sp. nov., but as no further details are available and the specimen cannot be found, we desist from assigning it species status and exclude the record from our map.
Dasypeltis atra Sternfeld
Montane Egg-eater
Figs 17A–M & 18
Dasypeltis lineolata Peters, 1878: 206. Type locality: Kitui, Kenya, holotype ZMB 9224.
Dasypeltis palmarum (not Leach) Fischer, 1884a: 12 (Arusha, Tanzania).
Dasypeltis scabra var. F, Boulenger, 1894: 356 (part, Uganda); Angel, 1925: 34 (Nairobi).
Dasypeltis palmarum ? (not Leach) Stejneger, 1894: 733 (Mount Kilimanjaro, Tanzania).
Dasypeltis scabra (not Linnaeus) Boulenger, 1896: 553 (Shoa, Ethiopia); Sternfeld, 1908: 240, 332 (Lake Haramaya, Ethiopia); Roux, 1910: 99 (Bukoba, Tanzania); Boulenger, 1909: 303 (Bugala, Sesse Islands, Uganda); Peracca, 1909: 173 (Fort Portal, Ruwenzori Mtns); Boulenger, 1911: 5 (Bussu, Uganda) & 1912: 332 (Addis Ababa); Loveridge, 1916: 79 (part, Fort Hall); Lönnberg, 1922: 7 (part, east slopes of Mount Elgon, Kenya); Loveridge, 1929: 28 (part); Parker, 1932: 220 (Lake George, Lake Naivasha); Loveridge, 1936a: 34 (part, Harar, Lake Bunyonyi) & 1936b: 256 (Uganda & Kenya); Mertens, 1938: 8 (Ngorogoro, Aberdares); Gans, 1959: 141 (part); Spawls, 1978: 10; Largen & Rasmussen, 1993: 340 (part); Flemming & Bates, 1999: 423 (Ibaya Camp, Mkomazi Game Reserve); Wagner & Böhme 2007: 136 (USNM 49376, Kakamega Forest).
Dasypeltis scabra var. atra Sternfeld, 1912: 272. Type locality: ‘Urwald hinter den Randbergen am Nordwestufer des Tanganyika-Sees’ [= Jungle behind the border mountains on the north-west shore of Lake Tanganyika], Democratic Republic of the Congo; Gans, 1961: 73.
Dasypeltis scabra var. palmarum (not Leach) Angel, 1925: 34 (Nairobi).
Dasypeltis scaber (not Linnaeus) Loveridge, 1929: 28 (part); Parker, 1932: 220 (Lake George, Lake Naivasha); Loveridge, 1936a: 34 (part, Harar, Lake Bunyonyi) & 1936b: 256 (Uganda & Kenya); Uthmöller, 1937: 113 (part, Moshi, Uru).
Dasypeltis scaber scaber (not Linnaeus) Bogert, 1940: 85 (part, Ganame, Ethiopia).
Dasypeltis scaber palmarum (not Leach) Loveridge, 1942: 285 (Uganda, Rwanda) & 1946: 106.
Dasypeltis scabra palmarum (not Leach) Loveridge, 1955: 11 (Gilo, South Sudan).
Dasypeltis scaber ater Laurent, 1956: 261; Curry-Lindahl, 1956: 47 (Rwanda); Hinkel, 1986: 25 (Rwanda).
Dasypeltis scabra atra De Witte, 1962: 139; Bourgeois, 1968: 284.
Dasypeltis atra Gans, 1964: 279, 1971: 461 & 1974: 47; Laurent, 1965: 14; Pitman, 1974: 119; Hinkel & Fischer, 1988: 16 (Rwanda); Hinkel & Fischer, 1990: 137 (Nyungwe Forest, Rwanda); Broadley & Howell, 1991: 32; Fischer & Hinkel, 1992: 144 (Nyungwe and Birunga Forests, Rwanda); Hinkel, 1986: 25 (Rwanda); Meirte, 1992: 57; Largen & Rasmussen, 1993: 340; Hughes, 1997: 68; Broadley, 1998: xxxii; Gravlund et al., 1998: 53 (Mbizi Forest Reserve, Tanzania); Necas & Modry, 1998: 22; Gravlund, 2001: 101; Vonesh, 2001: App. A, Table 3 (Kibale National Park, Uganda); Behangana & Goodman, 2002: 66 (Uganda); Spawls et al., 2002: 414; ? Spawls, 2004: 17 (Keriyo; may be referable to plain phase D. abyssina); Spawls et al., 2006: 129; Largen & Parker, 2004: 25; Wagner & Böhme 2007: 136 (Kakamega Forest); Gartner & Greene, 2008: 368; Bates & Broadley, 2008a,b (abstracts); Broadley & Bates, 2009: 50; Bates, 2009: 3; Largen & Spawls, 2010: 494 (part, ‘Mulu’ = Keriyo, may be plain phase D. abyssina); Bates et al., 2012: 28; Branch, 2015: 67; Göthel, 2015a: 19 & 2015h: 90; Van de Loock & Bates 2016: 16 (Ngangao Forest, Kenya); Mannel & Kucharzewski, 2017: 43; Spawls, 2017: 31 (Maasai Mara).
Dasypeltis scaber var. lineolata (in error) Trape & Mané, 2006: 53.
Description. There are three basic colour morphs: dorsum black (‘2M’ of Gans 1959) with a grey venter (N = 89); dorsum brown (‘2B’ of Gans 1959) with a cream, yellowish or pale orange (e.g. Fig. 17C) venter, usually with grey stippling at the edges (N = 90); dorsum grey or tan brown, decorated with closely-spaced, mostly narrow, transversely enlarged dark saddles, sometimes rhombic, in common or occasional contact with the equally dark lateral bars, or occasionally separated in a more-or-less ‘5N’ pattern similar to that of D. scabra (‘2Bx’ of Gans 1959) and venter cream, usually with grey stippling at the edges (N = 128). In patterned specimens the dorsal saddles (62–130, mean 91.8, N = 51) are dark brown, olive or reddish; and the head may be largely grey to bluish-grey, and/or with few dark markings unlike the boldly dark-light patterned head of D. scabra (see images in Broadley & Bates 2009; Spawls et al. 2002; Göthel 2015e,h; Fig. 17F, I–K). For example, PEM R17336 (Tanzania; Fig. 17K) and BM 03.12.2.22 (Uganda) have grey bodies with reddish saddles/crossbars; but the latter specimen also has a broad red chevron on the top of the head. Saddles and lateral bars may vary in colour between specimens even at the same locality – e.g. at Kitengela in Kenya they may be olive (Fig. 17F) or brown. The ‘lineolata’ phase (Gans, 1959, Pl. VI, Fig. 8; Fig. 17D) shows the extreme reduction of the pattern where most of the back is dark with only narrow, often interrupted or incomplete, pale transverse bars. Some specimens with low numbers of bands are somewhat similar to D. confusa, but lack the large well-defined saddles. Dorsally brown (‘2B’) and black (‘2M’) phase specimens are found together at a few localities such as Bourka in Ethiopia, Bussu, Kampala and Sipi at Mt Elgon in Uganda, and Minziro Forest Reserve in Tanzania; brown and patterned (‘2Bx’) phases are both found at Harar in Ethiopia, Mushongeru in Uganda, and Nairobi and Kacheliba in Kenya; black and patterned specimens both occur at Nakuru and North Nandi Forest in Kenya; and at Muko in Uganda all three colour phases occur together (Appendix 1). All specimens from South Sudan are melanistic (i.e. ‘2M’), except MCZ 53569 which is brown.
Preoculars usually one on either side of head (96.5% of specimens), but one on one side and two on the other in 1.8%, and two on either side in 1.8% (N = 227); postoculars one on either side of head (e.g. Fig. 18) in 58.3% of specimens, one on one side and two on the other in 6.8%, and two on either side in 34.9% (N = 307) (usually one in western populations – especially in Uganda, Rwanda and Burundi, e.g. 93.0% of Rwanda/Burundi snakes [N = 43] had a single postocular on either side; by contrast 77.4% of Ethiopian snakes [N = 31] had two postoculars on either side); frontal shield for the most part smooth, usually with 1–2 rows of shallow pits at the edges (States 1–2; 95.3% of specimens), but occasionally fully smooth (State 0; 3.3% of specimens) or extensively pitted (States 3–4; 1.4% of specimens: NMW 15299, PEM 16778, ZMUC 6122) (N = 213); inter-prefrontal suture usually weakly marked (87.5% of specimens), but moderately marked in 11.5%, and distinctly depressed in 1.0% (BM 1976.2265, MSNG 30412) (N = 200); supralabials 12–16, usually 14 (both sides of head), usually 7 [3rd and 4th entering orbit] on either side (84.7%), but variations are: 6[2,3] on either side of head (e.g. MRAC 18557) in 1.4% of specimens, 6[3,4] on either side (e.g. BM 51.1.3.74) in 5.2% (including MSNG 52410 which had left side of head damaged), 6[3,4] on left and 6[2,3] on right in ZFMK 29731, 6[2,3] on one side (e.g. CAS 122281) in 2.8%, 6[3,4] on one side (e.g. MCZ 40578) in 1.4%, 7[2,3,4] on right in ZFMK 16237, 7[2,3,4] on both sides in NMK 3182, 8[3,4] on one side (e.g. BM 1976.2265) in 1.0%, 8[3,4,5] on one side (e.g. USNM 40897) in 1.7%, and 8[3,4,5] on both sides (e.g. MRAC 15440) in 0.7% (N = 287); anterior temporals usually two on either side of head (93.9%), occasionally one (2.5%), 2 left:1 right or 1:2 (1.2%), 2:3 or 3:2 (1.2%), or three (1.2%) (N = 244); posterior temporals usually three on either side of head (52.7%), often two (18.3%) or 2:3 or 3:2 (14.1%), occasionally 3:4 or 4:3 (8.3%), four (4.1%) or five (0.8%), and exceptionally 4:5 (ZFMK 29733), 3:5 (BM 1976.2265), 5:3 (BM 34.12.15.590) or 2:4 (LIV 1960.67.1) (N = 241); temporal formula on either side of head usually 2+3, often 2+2 or 2+4; midbody scale rows 22–28 (mean 24.1 ±1.25, N = 226) [28 in NMW 26879/3 only]; ventrals 199–232 in males (N = 109), 211–256 in females (N = 167); subcaudals 50–71 in males (N = 103), 45–64 in females (N = 151). One unsexed specimen from Bujumbura, Burundi (MRAC 82–16–R4, probably a male) had 218 ventrals and as many as 75 subcaudals. The brown (‘2B’ of Gans 1959) female (MSNM 7202) from Circao (northern-most record for this species) has 232 ventrals, a much lower count than that of female D. abyssina (241–264) which also occur as brown or uniform plain phase morphs (see below); and brown D. atra in Ethiopia have lower ventral + subcaudal counts than plain phase D. abyssina (both sexes, 269–291 [N = 5] versus 294–323 [N = 7] respectively). Ratio SVL: tail length 4.2–6.7 in males (N = 97), 5.8–8.3 in females (N = 143); total length: tail length 5.2–7.7 in males (N = 97), 6.8–9.3 in females (N = 143).
INSERT FIGS 17 & 18
Size. Largest male (MSNG 30413b – Bussu, Uganda) 611 + 98 = 709 mm; largest female (NMK 2583 – North Nandi Forest, Kenya) 890 + 115 = 1005 mm.
Habitat. Found mainly in high elevation savannah and forest, usually above 1500 m.
Distribution. Kivu Region and vicinity of Lake Albert in eastern Democratic Republic of the Congo (Laurent 1956; Gans 1959), Rwanda, Burundi, south to Mbizi Forest in western Tanzania and east to the extreme northern parts of that country, Uganda, south-western, central and southern Kenya, extreme southern part of South Sudan, south-western and central Ethiopia, with the northern-most record at Circao in north-western Ethiopia (Fig. 3).
Sympatry. Sympatric with D. medici at Nyambene Hills (Kenya) and Moshi (Tanzania); sympatric with D. confusa 10 km S of Juba (South Sudan), at Busingiro and Gulu (Uganda) and Kacheliba (Kenya); sympatric with D. scabra at Aware Melca and Lake Abiata (Ethiopia), Mulema (Uganda), Lake Naivasha and Mount Kulal (Kenya), Astrida (Rwanda) and Minziro Forest Reserve (Tanzania); sympatric with both D. medici and D. scabra at Mount Kilimanjaro (Appendix 1).
Localities. ETHIOPIA. ‘Abessinien’, NMW 9964/2; Addis Ababa [NE 0938B2] MSNG 30412, and Fig. 17E; Aware Melca [NE 0939B2] BM 16.6.24.13; Bedele [NE 0836A4] ZFMK 57537; Bedele, 4.5 km SW of [NE 0836A4] melanistic, photographic records; Bourka [NE 0941A4] MNHN 05.208–10; Circao, Dambria [NE 1237A4] MSNM 7202; El Dire, Tertale [NE 0437C3] MSNM 2975; Fitu, 135 km ESE of Negele [NE 0540B1] ‘2Bx’ pattern, photographic records; Ganame [NE 0841D3] AMNH 20345; Gemu, Gofa [NE 0636B] BM 1964.1068; Genale to Chamo [NE 0837D3] ZFMK 16297; Ghimbi, Wollega [NE 0935B2] BM 1973.3175; Ginir [NE 0740B1] NMW 26879/2; Gololtscha [NE 0840A1] ZFMK 5535; Gumaydo [NE 0537D1] NMW 26879/3; Harar [NE 0942A3] FMNH 4012, MCZ 34923, NMW 9964/1; Jigjiga [NE 0942B4] ZFMK 84993; Kebre Mengist to Negele [NE 0539C3] BM 1975.2144; Keriyo [NE 0938B3] (Fig. 17A); near Kersa Dek, 14 km SE of Neghelle [NE 0539B1] ‘2Bx’ pattern, photographic record; Kounni [NE0940B2] MNHN 05.211; Laga Hardine [NE 0840C4] MNHN 05.205–7; Lake Abiata [NE 0738D1] ZFMK 16236; Lake Alemeya [NE 0942A3] ZMB 27463; Lake Shala [NE 0738B3] ZFMK 16237; Let Marefia [NE 0939D2] ZMB 10286; Neghelle [0539B3] (Fig. 9M); Shek Husen, 4.4 km WSW of [NE0740D1] brown dorsally with cream patches, photographic record; Shoa, MSNG 29047a; Wondo [NE 0638C2] NMW 15299. SOUTH SUDAN. Gilo, Imatong Mtns [NE 0432B2] BM 1978.379–80, FMNH 62215, MCZ 53406–8, 53569, ZFMK 29564, 29719, 34532, 59567; 10 km S of Juba [NE 0431D3] ZFMK 29567; Katire [NE 0432B2] ZFMK 26005; Talanga Forest [NE 0432B1] ZFMK 29731–3. UGANDA. Bihunga [SE 0129B4] LIV 1960.67.1, 1960.50.32; Bitereko [SE 0030B3] LIV 1960.115; Budongo Forest [NE 0131D3] USNM 63481; Bufindi, Kigezi [NE 0129B4] BM 51.1.3.68 & 79, NMZB 217; Bukakata [SE 0031A3] BM 51.1.3.80; Busingiro, Budango [NE 0131D1] BM 1951.1.5.24; Bussu [NE 0033A3] BM 11.7.8.14, MSNG 30413a, b, 52410; Butundiga, Mount Elgon [NE 0134A2] MCZ 40574; Bwamba Forest [NE 0030C3] LACM 38696, 46344; Chagwe, Mabira Forest [NE 0032D2] BM 08.10.20.14–5; Entebbe [NE 0032A2] BM 51.1.3.74; Gulu [NE 0232C4] BM 51.1.3.77; Fort Portal [NE 0030C2] CAS–SU21709–10; Kabingo, Bunyaruguru, BM 1961.1803; Kagabagara [SE 0030D3] LIV 1962.9.28; Kagando [NE 0029B2] LIV 1960.67.5; Kalinzu Forest [NE 0030A3] LACM 60705; Kampala [NE 0032A2] BM 36.7.3.32, 1984.882; Katunguru [SE 0030A1] NMZB 103; Katwe [SE 0029B2] MRAC 9997; Kayonza, Kinkizi [NE 0029D3] LACM 35090, LIV 1960.115.4, 1962.9.27, 1980.150.31, NMZB–UM 2550; Kazinga Channel [SE 0030A1] NMK 404; Kichwamba [SE 0030A1] LIV 1962.9.14; Kinani [SE 0030B4] LIV 1962.9.22; Kyamahunga, Igara [SE 0030A3] BM 51.1.3.75, 1961.1805, NMZB 2549; Kyamugorani [SE 0030B3] LIV 1962.9.4; Lake Bunyonyi [SE 0129B4] FMNH 9913–4, MCZ 34922; Lake Edward [SE 0029B] NMZB 219; Lake George [NE 0030A] BM 32.5.2.99; Lake Nabugabo [SE 0032A3] BM 51.1.3.78; Likiri [SE 0030A2] LIV 1980.150.27; Maozi-Isingiro [SE 0030D4] BM 03.12.2.23; Mihunga, Ruwenzori [NE 0030A3] MCZ 48359; Mityana [NE 0032A3] NMK 3291; Morulinga, 24 km SW of [NE 0234A4] BM 1954.1.12.37; Mubango, Mabira Forest [NE 0033A3] BM 34.12.15.592; Muko, Kigezi [SE 0129B2] BM 34.12.15.585–91, 593–4; Mulema [SE 0031D1] BM 03–12–2–22; Mushongero, Lake Mutanda [SE 0129B1] MCZ 48362–3; Nsika [SE 0030A4] LIV 1980.150.26; Nyabushozi [SE 0030B4] LIV 1962.9.23; Nyakabende [SE 0129B4] MCZ 48360–1, UMMZ 88532/1; Nyamuhumia [SE 0030C1] LIV 1959.81.2; Nyansimbo [SE 0030B1] LIV 1962.50.33, 1980.150.33; Nyarusiza, Bufumbira [SE 0129B3] BM 1976.1849, LIV 1980.150.29–30, NMK 1496, NMZB–UM 2553; Rucho, Ankole, MCZ 54744; Ruhama [SE 0030C4] LIV 1960.67.3; Sango Bay Forest [SE 0031D3] LACM 39242; Sesse Islands, Lake Victoria [SE 0032A3] MSNG 27755a, 40764; Sipi, Mount Elgon [NE 0134A4] MCZ 40564–73. KENYA. No locality, NMK 348, 463, 1962, UMMZ 96274; Athi Plain, NMW 9964/20; Athi-Kapiti Plains, NMK 3129; Athi River [SE 0136B4] NMK 1453, 3085; Butere, Kakamega [NE 0034B1] NMK 2323 (old no. 306); Chemelil, Kisumu [SE 0035A1] CAS 148034–5; Chuka, Meru [SE 0037B3] CAS 122281, and Fig. 17G – ‘2Bx’ pattern; Chyulu Hills [SE 0037D2] NMK 406; Elgonyi, Mount Elgon [NE 0134B1] MCZ 40575; Emakoko, Nairobi National Park [SE 0136B4] photographic record (dorsum grey with brown saddles [similar to Fig. 9F] and two postoculars on left side of head); Embakasi, Nairobi [SE 0136B4] NMK 2849; Hopcroft Ranch, Athi River [SE 0137C1] NMK 3131; Juja Farm [SE 0137A1] USNM 40897; Kabartonjo, Kamasia Mtns [NE 0035D2] CAS 111767; Kabarnet, Baringo [NE 0035D2] BM 1957.1.11.42, NMK 3329; Kabete, Kiambu [SE 0136B3] NMK 479; Kacheliba [NE 0135A3] NMK 486, 488; Kaimosi [NE 0034B2] MCZ 40576–8, 18205, UMMZ 61224, USNM 49078; Kairuni, Nyambeni Mtns [NE 0037B2] LACM 93378–9; Kakamega [NE 0034B4] BM 1959.1.5.58, CAS 142248, NMK 482, 2715, USNM 49376; Kakamega Forest [NE 0034B4] Fig. 16L, ‘2Bx’ pattern; Karen, Nairobi [SE 0136B3] NMK 2503, 3007; Kericho [SE 0035A4] NMK 1260; Kiambu [SE 0136B2] NMK 481; Kiboko Ranch Station [SE 0237B1] NMK 2952 (4); Kijabe [SE 0036D3] NMK 2130, and top photograph of melanistic specimen in Spawls et al. 2002: 415; Kiganjo, Nyeri [SE 0036D4] NMK 2862; Kiserien, Baringo [SE 0136B3] NMK 3123; Kitale [NE 0135A1] NMK 3051, ZMUC 6101; Kitengela [SE 0136D2] Fig. 17F and another photograph, both ‘2Bx’ patterned; Kitengela, Athi River [SE 0136D2] NMK 3413; Kitui [SE 0138A3] ZMB 9224 (holotype of D. lineolata); Lake Baringo [NE 0036C2] NMK 1791, 4174; Lake Naivasha [SE 0036C2] BM 93.11.21.72, 1932.5.2.100; Limuru [SE 0136B1] Fig. 17B, melanistic; Lorgorien, Trans-Mara, NMK 3109; Lumbwa, Pen-y-Bryn [SE 0035A4] CM 2278–9, 2281; 13 km SSW of Maralal [NE 0036D3] LACM 66076; Mbunyi [SE 0137B4] USNM 62907 (Gans’ 1959 locality of ‘Morogoro’ for this specimen is in error, see Loveridge 1929); Mount Elgon, NMK 657–9, 3152; Mount Kenya [SE 0037A2] USNM 40990; Mount Kulal [NE 0236D2] LACM 66395; Muranga [Fort Hall] [SE 0037C1] BM 08.9.17.4; Mweiga [SE 0036B4] LACM 50631; Mwino, Sigor [NE 0135A4] NMK 1965; Nairobi [SE 0136B4] NMK 403, 2486, 3339, 3548; Naitolia (6000 ft = 1829 m a.s.l.) [SE 0036A4] BM 01.1.3.9; Nakuru [SE 0036A3] NMK 484 and 1453, and photographs of ‘2Bx’ patterned and melanistic specimens; Nandi [NE 0035A1] BM 06.7.6.7; Nandi Hills [NE 0035A] NMK 1158; Nanyuki, Laikipia [NE 0037A1] NMK 3490, 3512, 3557; Nbabibi, Naivasha [SE 0036C2] NMK 1960; Ngangao Forest, Taita Hills [SE 0338A4] (photograph of barred/patterned ‘2Bx’ specimen: Van de Loock & Bates 2016, Fig. 2); Ngong [SE 0136B3] BM 1960.1.6.40; North Nandi Forest, Chemesia [NE 0034B4] NMK 2576, 2583, 2590; Nyambene Hills [NE 0037B] NMK 1487, 1489, 1699; South Nandi Forest [NE 0035A1] NMK 4176; Subukia [SE 0036A1] FMNH 58337; Timbilil, Mau Forest [SE 0035A4] NMK 3182; Uganda Railway, BM 96.3.27.18. RWANDA. Astrida [SE 0229D1] MRAC 18554–8; Bugarama [SE 0229C1] MRAC 86–01–R–10 & 11; Bugesera [SE 0230A1] ZFMK 61616; Bulimbi [SE 0130C1] MRAC 18577; Cyamudongo [SE 0228D2] ZFMK 61617; between Gikongoro and Nyungwe Forest Reserve [SE0229C2] PEM 5623; Kamohora District, MRAC 14329; Kibungo [SE 0230B1] ZFMK 55105; Kisenyi [SE 0129C1] MCZ 48364, MRAC 2723, 2725, 2757, 3821, 5871–2, 9773, ZMB 20458, 27156, 27539; Lake Gando [SE 0129C2] MRAC 14333, 14341; Mwogo River near Butare [0230A1] MRAC 74–23–R–22; Mutura, Mount Virunga [SE 0129B2] MRAC 7509; Nyanza [SE 0429B3] MRAC 15477; Ruhengeri [SE 0129D1] IRSNB 2657; Wankeri [SE 0129D1] MRAC 14331, 14340: Rwankuba [SE 0129C2] MRAC 18259; Shangugu [SE 0228B4] MRAC 4074–5; Tare, Astrida [SE 0229D1] MRAC 18560–2. BURUNDI. Bujumbura [SE 0329A4] MRAC 82–16–R3 & 4; Kamagaba Marsh, MRAC 18583; Kibira Forest [SE 0229C4] (Fig. 17C); Makamba [SE 0429B2] MRAC 18584; Rumonge [SE 0329C4] MRAC 18585. TANZANIA. Bukoba [SE 0131B4] ZMB 16749–50; Ibaya Camp, Mkomazi Game Reserve [SE 0337D4] BM 1994.403, NMBO 7375; Kasoge, Mahale Peninsula [SE 0629B2] BM 1970.2177; Klein’s Camp, NE Serengeti National Park [SE 0135C3] PEM 16778, 17329, 17336, 17941; Masiko ya Tembo, vicinity of Sanya Chini [SE 0337A3] (Fig. 17J); Mbizi Forest Reserve [SE 0731D3] ZMUC 6122; Minziro Forest Reserve [SE 0131B1] ZMUC 6138–41; Moru area, Serengeti National Park [SE 0234D1] photograph of ‘2Bx’ marked specimen; Moshi [SE 0337A4] ZSM 81/1936 (3); Mount Kilimanjaro [SE 0337A] ZMB 17039 (2); Oldeani [SE 0335B3] photograph of melanistic ‘2M’ specimen; Sitete, Mahale Peninsula [SE 0629B2] BM 1970.2178.
Conservation. In terms of the regional (study area) and global distribution of this species, using IUCN (2012, 2016) criteria, it is widespread and abundant and should be regarded as Least Concern.
Remarks. Dasypeltis scabra var. atra was described by Sternfeld (1912) on the basis of three ‘completely black’ specimens, with single postoculars, from the forest behind the border mountains on the north-west shore of Lake Tanganyika, Democratic Republic of the Congo. Photographs of the three syntypes housed at Museum für Naturkunde, Berlin were provided by F. Tillack (pers. comm, 10 July 2017). ZMB 27761 is medium brown above and light brown below (may have faded); ZMB 22516A and ZMB 22516B are dark brown to black above and grey below. It can be confirmed that all three specimens have a single postocular on either side of the head, and the supraocular extends over the posterior corner of the eye.
Since its description by Peters in 1878, Dasypeltis lineolata was used as a valid name by Fischer (1884b) in reference to a snake (247 ventrals, 69 subcaudals) from Ssibange in Gabon, probably referable to D. fasciata or possibly D. palmarum, and later by Stejneger (1894) who concluded that it was a ‘colour variety’ (possibly implying a subspecies) of Dasypeltis abyssina. The only other valid usage of the name was by Saleh & Sarhan (2016). While referring to available names, Trape & Mané (2006) erroneously used the combination Dasypeltis scaber var. lineolata. The samples of ‘D. cf. lineolata’ from Burundi used in the 16S genetic analysis of Trape et al. (2012) are referable to either D. atra or D. scabra. The name D. atra (or D. scaber ater or D. scabra atra, as indicated), the junior synonym, has been used in the following 42 publications (25 required), by 34 authors/sets of authors (10 required), during the preceding 50 years (1968–2017), and for a period covering more than 10 years (see Article 23.9.1.2 of the ICZN 1999): Bourgeois (1968, as D. scaber ater and D. scabra atra), Gans (1971, 1974), Pitman (1974), De Witte (1975), Hinkel (1986, as D. scaber ater), Hinkel & Fischer (1988, 1990), Fischer & Hinkel (1992), Broadley & Howell (1991), Meirte (1992), Largen & Rasmussen (1993), Hughes (1997), Broadley (1998), Gravlund et al. (1998), Neças & Modry (1998a,b), Gravlund (2001), Spawls et al. (2002), Largen & Parker (2004), Spawls (2004), Branch (2015), Spawls et al. (2006), Trape & Mané (2006), Wagner & Böhme (2007), Gartner & Greene (2008), Bates (2009), Broadley & Bates (2009), Largen & Spawls (2010), Bates et al. (2012), Trape et al. (2012), Göthel (2015a–h), Van de Loock & Bates (2016), Saleh & Sarhan (2016), and Mannel & Kucharzewski (2017), thus satisfying the requirements of Article 23.9.1.2 of the ICZN (1999). However, Saleh & Sarwan’s use of the name D. lineolata means that the provisions of Article 23.9.1.1 (i.e. senior synonym not used as a valid name after 1899; ICZN 1999) are not met. In order to maintain nomenclatural stability the ICZN has been requested (Bates 2017, submitted; Case 3750) to suppress the older name D. lineolata by taking action according to Article 23.9.3 by use of its plenary power (Article 81; ICZN 1999). We maintain prevailing usage of the name D. atra according to Article 82.1 (ICZN 1999).
Dasypeltis fasciata A. Smith
Western Forest Egg-eater
Fig. 19A–C
Dasypeltis fasciatus A. Smith, 1849 (footnote to text accompanying Plate 73). Type locality: Sierra Leone.
Dasypeltis scaber fasciatus Loveridge, 1942: 284 (Bundibugyo, Uganda).
Dasypeltis fasciata Loveridge, 1957: 289 (check-list); Gans, 1959: 137 & 1961: 73; Pitman, 1974: 121; Spawls et al., 2002: 416; Spawls et al., 2006: 130; Branch, 2015: 68; Göthel, 2015a: 19 & 2015h: 81.
Description. Dorsum grey or pale brown with numerous broad dark saddle bands (108–141, N = 5) which are often ill-defined (preserved specimens, e.g. Fig. 19A) and usually arranged in a manner similar to the ‘5N’ pattern, with saddles alternating with broad lateral bars; dorsal body pattern expressed as chequerboard on skin between scales and on lower edges of these (see Gans 1959, Pl. III); top of head vermiculate; venter cream to pale grey, usually with grey stippling at the edges, but sometimes extensively stippled (e.g. TCWC 23839, which also has the throat and anterior part of the belly pale yellow, Fig. 19B).
Preoculars usually one on either side of head (83.3% of specimens), but two on either side in MCZ 48357 and UMMZ 88531 (N = 12); postoculars usually two on either side of head, but two on left side and one on right side of MCZ 48358B, and one on both sides in LACM 46341 (N = 12); frontal shield for the most part smooth, usually with 1–2 rows of shallow pits at the edges (States 1–2), but more extensively pitted (State 3) in LACM 38697 (N = 11); inter-prefrontal suture weakly marked (N = 11); supralabials 13–14, usually 14 (both sides of head), usually 7 [3rd and 4th enter orbit] on either side of head, but 6 [2,3] on right side of head in LACM 46346 and MCZ 48357 (N = 12); anterior temporals usually two on either side of head (75.0%), occasionally 2 left: 3 right or 3:2 (16.7%), and three in LACM 46341 (N = 12); posterior temporals usually three on either side of head (58.3%), occasionally 3:4 or 4:3 (25.0%), two in LACM 39243, and four in LACM 46343 (N = 12); temporal formula on either side of head usually 2+3, occasionally 2+2, 2+4, 3+3 and 3+4; midbody scale rows 22–25 (mean 23.5 ±0.80, N = 12); ventrals 212 (LACM 39243), 229–230 in males (N = 3), 237–248 in females (N = 9); subcaudals 59 (LACM 39243), 71–76 in males (N = 3), 64–76 in females (N = 9). Ratio SVL: tail length 4.7–5.2 in males (N = 3), 4.8–6.4 in females (N = 9); total length: tail length 5.7–6.2 in males (N = 3), 5.8–7.4 in females (N = 9).
INSERT FIG. 19
Size. Largest male (LACM 38697 – Ntandi, Bwamba Forest, Uganda) 525 + 111 = 636 mm; largest female (TCWC 23839 – Ntandi, Bwamba Forest, Uganda) 611+ 108 = 719 mm.
Habitat. Lowland forest at elevations of about 1000 m to 1147 m (Katera).
Distribution. Forested areas of west and central Africa, from Guinea to Togo, and from south-eastern Nigeria south to north-western Angola, and east through southern Central African Republic and much of the Democratic Republic of the Congo (Gans 1959, 1964; Trape & Mané 2012), reaching its eastern limit in western and southern Uganda (Fig. 3).
Localities. UGANDA. Bundibugyo [NE 0030C] MCZ 48537, 48538a,b; UMMZ 88531; Bwamba Forest, Ntandi [NE 0030C3] LACM 38697, 46341–3, 48345–6, TCWC 23839; Katera, Sango Bay Forest [SE 0031D3] LACM 39243.
Conservation. In terms of the regional (study area) and global distribution of this species, using IUCN (2012, 2016) criteria, it appears to be widespread and abundant and should be regarded as Least Concern.
Dasypeltis abyssina (Duméril, Bibron & Duméril)
Ethiopian Egg-eater
Figs 20A–G
Rachiodon abyssinus Duméril, Bibron & Duméril, 1854: Vol. 7: 496, Pl. 81, Figs 1–2. Type locality: Abyssinia, proposed lectotype MNHN 6567 (see below); Duméril, 1860: 198.
Rachiodon scaber var. abissinicus (lapsus) Jan, 1863: 106 (Ethiopia).
Dasypeltis abyssinicus (lapsus) Blanford, 1870: 457 (Senafe, Eritrea); Zavattari, 1930: 193.
Dasypeltis abyssina Fischer, 1884b: 48; Trape & Mané, 2006: 53; Largen & Spawls, 2010: 490; Göthel, 2015a: 19, 2015h: 92.
Dasypeltis scabra (not Linnaeus) Boulenger, 1893: 354 (var. D, Abyssinia); Calabresi, 1925: 107 (Gondar); Zavattari, 1930: 193 (Senafe); Gans, 1959: 141 (part, Eritrea, Gheleb; Pl. VI, Figs 3 &4); Largen & Rasmussen, 1993: 340 (part, 140 km NE of Gondar on road to Asmara); Spawls, 2004: 17 (Keriyo); Largen & Spawls, 2010: 488 + 490 (part, e.g. ‘Mulu’ = Keriyo).
Dasypeltis scaber (not Linnaeus) Loveridge, 1936: 34 (part, Gondar).
Lectotype. Dumeril et al. (1854) described Rachiodon abyssinus on the basis of two syntypes in the Muséum national d’Histoire naturelle in Paris, one from “Abyssinia” (= Ethiopia) and another collected during an expedition to the sources of the White Nile under direction of M. Arnaud (labelled “Nil Blanc” = White Nile, perhaps above Khartoum in Sudan). Although their description is compound (e.g. ventrals are given as 222–260, clearly referring to MNHN 7227 and MNHN 6567 respectively – see above), the information on colour, which refers to a yellow base colour which exaggerates the darker dorsal markings, appears to be specifically in reference to the illustrated snake (MNHN 6567). Trape & Mané (2006) re-instated D. abyssina which has vivid ‘5N’ dorsal colouration, heavily black-bordered ventrals and a high ventral count of 260. They were of the opinion that the two syntypes were probably referable to different species, and designated MNHN 6567 (female), illustrated in plate 81 in the description (see also Fig. 20A–B), as ‘holotype’ of R. abyssinus. Although we agree that the two specimens do in fact represent different species, the action by these authors is not in accordance with the International Code on Zoological Nomenclature (ICZN 1999) as a syntype cannot be designated as a holotype, only as a lectotype or paralectotype.
MNHN 6567 agrees closely with the heavily-marked belly illustrated in plate 81 of Duméril et al. (1854), but the dorsal pattern is dissimilar (Fig. 20A). The illustration depicts a snake with a pale vertebral stripe, flanked on either side by distinct but disjointed markings. As can be seen in the photograph (Fig. 20A) of this specimen, while the centre of the back has a pale background, this is interrupted frequently by the often oblique dark markings or saddles which are continuous across the centre of the back. There is also a general separation of the saddles from the lateral bars, even though these may be in occasional contact. An indication of the cause of the poor condition of the specimen (head and neck damaged) is provided by plate 81 in which the snake is illustrated, together with a section of its skeleton comprising 13 pairs of ribs and their associated vertebrae, and a section of the vertebrae from the neck region, indicating considerable mutilation of the body. It was therefore not possible to perform a count of the ventral plates, and as an appreciable portion of the tail tip was missing, no subcaudal count was possible either. Nevertheless, there seems to be no doubt that the snake illustrated in plate 81 and described by Duméril et al. (1854) is the specimen represented by MNHN 6567.
We take this opportunity to designate MNHN 6567 (female, no hemipenal spines in tail) as lectotype of Rachiodon abyssinus. In accordance with Recommendation 74C of the 1999 Code we hereby list the following data pertaining to the lectotype, as based on a recent examination of the damaged specimen (Fig. 20A–B) in Paris by the first author: Type locality: “Abyssinia” (= Ethiopia). Snout-vent length about 572 mm; about 86 pattern cycles from nape to above vent. Dorsal colour pattern consists mostly of oblique dark saddles, usually wider than long, and often confluent or forming zigzags on the back and tail; lateral bars (usually extending onto the edges of the ventral plates) somewhat irregular, usually broader above than below, and often in contact with the pale interspaces between saddles, sometimes in narrow contact with the edges of the saddles, but never forming a distinct or broadly linked pattern. Ventral scales with extensive brown markings on their anterior half, consisting of a large rounded cone on either side joined to a smaller cone in the centre. Midbody scale rows 25; lateral dorsal scale rows 3 to 6 are keeled and serrated; nasal scale divided between nostril and first upper labial.
Paralectotype. Duméril et al.’s (1854) other syntype of R. abyssinus, MNHN 7227 (female, no hemipenal spines in tail) from ‘Nil Blanc’, now the paralectotype, is referable to D. scabra. It measures 673 + 90 mm and has a grey dorsum with ‘5N’ pattern (55 pattern cycles) consisting of brown oval saddles with black borders, and lateral bars—situated adjacent to the pale inter-saddle bars—also brown with black borders (Fig. 6); venter cream with dark speckling and two longitudinal rows of large spots at the edges; preoculars 1 on either side of head; postoculars 2 on either side; frontal shield with marginal pitting (State 2); inter-prefrontal suture moderately developed; ventrals 226; subcaudals 52; midbody scale rows 25, rows 3–5 keeled and serrated; supralabials 7 (3rd and 4th enter orbit) on either side; nasal scale divided below the nostril; temporals 2+3.
Diagnosis. Assigned to the genus Dasypeltis on account of its slender form, possession of (usually 3–4) rows of reduced, oblique, keeled and serrated lateral scales (little or no serration in D. inornata) and head barely distinct from the neck (moderately distinct in D. fasciata). Occurs as both patterned and uniform (brown or grey above, immaculate cream below) phases. Patterned individuals are distinguished from other species by their unique dorsal and ventral colour patterns, dorsally consisting mostly of oblique dark saddles, usually wider than long, and often confluent or forming zigzags on the back and tail; lateral bars much-elongated, usually broader above than below, and typically extending onto the edges of the ventral plates, irregular in shape and often in contact with the pale interspaces between saddles, sometimes in narrow contact with the edges of the saddles; pattern cycles 65–98 (≤62 in D. crucifera sp. nov., D. taylori sp. nov. and D. bazi; D. inornatus always uniform brown above; D. gansi uniform or weakly marked above); and ventral scales with extensive black, grey or brown markings on their anterior half, consisting of a large rounded cone on either side joined to a smaller rounded cone in the centre (Gans 1959, Pl. 9, Fig. 7; Fig. 20B; some D. scabra have heavy dark markings on the venter, but never as extensive or in the same pattern as described above); in live specimens the labials and interspaces between saddles are cream-yellow (Fig. 20C–F) as illustrated for one of the two syntypes (MNHN 6567, see discussion below) which, in addition, apparently also had some yellow on the sides of the head (Duméril et al. 1854, Pl. 81): within the genus this combination of dark markings with extensive cream-yellow or yellow also occurs, occasionally, only in southern populations of D. scabra (e.g. Peters 1864; Göthel 2015). In addition to its colour pattern as described above, this species can also be distinguished by the following combination of characters: High ventral scale counts of 226–247 in males (higher or mostly higher than D. scabra 180–226, D. taylori sp. nov. 196–216, D. confusa 199–227, D. sahelensis 207–221, D. inornata 208–225, D. parascabra 210–223), 241–271 in females (higher or mostly higher than D. scabra 198–249, D. taylori sp. nov. 208–222, D. sahelensis 212–237, D. confusa 212–242, D. bazi 215–232, D. inornata 219–237, D. parascabra 224-237); Subcaudals 57–68 in males (mostly higher than D. loveridgei comb. nov. 47–57; lower or mostly lower than D. inornata 81–92, D. gansi 68–83, D. palmarum 68–77, D. parascabra 67–74, D. latericia 66–86, D. fasciata 59, 71–91, D. medici 69–109), 49–65 in females (mostly lower than D. inornata 69–84, D. fasciata 64–84, D. medici 61–90); Inter-prefrontal sulcus usually weakly marked (weakly to moderately marked in D. bazi, usually moderately marked in D. taylori sp. nov., moderate to well marked in D. arabica sp. nov., and variable in D. scabra [often well marked in the south of its range]); Frontal marginally pitted, usually State 1 or 2 (extensively pitted, States 3 & 4, in D. medici, D. taylori sp. nov. and many populations of D. scabra especially in the south of its range); Tail of moderate length: SVL/tail length 5.3–6.1 in males, 5.5–8.1 in females (in study area mostly greater than: D. medici 3.5–4.7 males, 4.5–5.7 females; D. fasciata 4.7–5.2 males, 4.8–6.4 females; see also Gans 1959); Nasal divided below the nostril (undivided in D. sahelensis and D. parascabra); Postoculars 2 on either side of head (often one, especially in western populations, of D. atra); Supralabials usually 7 (3rd and 4th enter orbit) on either side of head (usually 6[2,3] in D. loveridgei comb. nov.).
Ventral counts in D. abyssina are mostly higher (226–247 in males, 241–271 in females) than the nearest populations of D. scabra in south-eastern Sudan and adjacent South Sudan (213; 222–227) and Ethiopia (212–218; 219–243), and much higher than D. taylori sp. nov. which occurs in northern Somalia and Djibouti (196–216; 208–222) (Appendix 1). Dorsally plain brown specimens are similar to the brown/grey phases of D. atra in north-eastern Africa and D. scabra in South Africa (e.g. De Waal 1978), although the venter may be yellowish-cream (MCZ 80984) rather than the usual plain cream. In addition, D. abyssina can often be separated from D. atra on account of its higher ventral counts (199–232 in atra males, 212–256 in females) and possession of two (rather than 1–2) postoculars on either side of the head. Plain phase D. abyssina are always distinguished from plain phase D. atra in Ethiopia (the only area where the two species are sympatric) by having higher ventral + subcaudal counts (both sexes, 294–323 [N =7] versus 269–291 [N = 5] respectively).
Description. Dorsum of patterned specimens with oblique dark saddles (65–98, N = 11), usually wider than long, and often confluent or forming zigzags on the back and tail. Lateral bars dark, long, mostly broader above than below, somewhat irregular and often in contact with the pale interspaces between saddles, sometimes in narrow contact with the edges of the saddles, and may extend onto the ventral plates as in the holotype, but never forming a distinct or broadly linked pattern. Ventral scales cream with extensive black, grey or brown markings on the anterior half, consisting of a large rounded cone on either side joined to a smaller cone in the centre. In live specimens the labials and interspaces between saddles are cream-yellow, and there may also be some cream-yellow on the sides of the head.
Four specimens from Eritrea in the Vienna collection (NMW) were listed under D. scabra by Gans (1959), with the dorsal patterns of three of them recorded as ‘5I’ (i.e. black rectangular saddles and lateral bars forming complete closure around the pale inter-saddle areas; see Gans 1959, Pl. VI, Fig. 4, and Fig. 20G). This material was examined in detail by the first author and all four specimens have at least some indication of the ‘5I’ D. arabica sp. nov.-like dorsal pattern: NMW 15300 has almost the entire dorsum patterned ‘5I’; NMW 15301–3 is similar but has mostly D. abyssina-like markings (as described above) at midbody; NMW 15301–1 has mostly abyssina-like markings anteriorly and mostly ‘5I’ markings posteriorly, although the pattern is not synchronised on either side of the midline just posterior to midbody (Gans 1959, Pl. VI, Fig. 3); while NMW 15301–2 has abyssina-like markings on the first quarter of the body and ‘5I’ markings on the rest of the dorsum. Gans (1959: 125) noted that although some ‘5I’ specimens had an immaculate venter (presumably including ZMB 7631 from Bogos = D. crucifera sp. nov. and BM 03.6.21.41 from El Kubar = D. arabica sp. nov. – see below), NMW 15301–2 (Gans, 1959, Pl. IX, Fig. 7) and NMW 15301–3 both had heavily dark-bordered ventrals like the holotype of D. abyssina (Duméril, Bibron & Duméril 1854, Pl. 81, Fig. 1–2; Fig. 20B). Re-examination of this material indicated that NMW 15301–1 also has the typically well marked abyssina-like ventral pattern, although the markings are pale grey in colour, while the ventral pattern of NMW 15301–3 is actually somewhat different to the others in that it consists of a large bold dark spot at the edges of the ventral plates, usually with 2–3 more-or-less disconnected paler blotches towards the centre. The distinct ventral markings of NMW 15300 are brown, which is perhaps an artifact of preservation.
INSERT FIG. 20
A plain tan brown to greyish phase with a largely unmarked cream to cream-yellow venter is also known, sometimes occurring in sympatry (at Gondar and Keriyo) with the patterned phase. In fact one of the six males and as many as six of the nine females are plain phase (marked by asterisks in ‘Localities’ below). A detailed description of two plain phase specimens follows: MCZ 80984 (Gondar to Asmara): dorsal scales are mostly pale tan, but the edges at the base and sides (anterior proximal half) of each scale is grey; NMB R10257 (Keriyo): dorsal scales pale tan with numerous tiny dark dots, and basal one-third of scale, as well as area around apical pits, grey. In both specimens the lowermost scale row on either side is without dark markings, while the scales of the adjacent row above have few dark markings on the lower edges of the scales, but the upper part of the scale base is dark; dark markings at the base of scales creates the impression of a very weak vertebral stripe and similar lateral bands, especially in NMB R10257. Dorsal scales of the latter specimen (and the plain phased NMZB 17099, also from Keriyo) are more elongate than in patterned conspecifics, and have rounded tips (compared to dorsals less elongate, sharply triangular and acutely-tipped in NMB R10256, a patterned specimen from the same locality as NMB R10257). However, the shape and serration of dorsal scales in MCZ 80984 is similar to that of patterned specimens. Belly and subcaudal region immaculate cream. In addition, these three plain phase specimens have only two rows of weakly serrated, inclined and reduced dorsals on either side of the body.
Preoculars 1 on either side of head (N = 18), postoculars usually 2 on either side of head, but one in BM 1902.12.13.86 (N = 18); frontal shield usually for the most part smooth with marginal pitting (States 1 & 2; with a single row in most specimens examined in detail), but entirely smooth in BM 1902.12.13.85 (State 0) (N = 13); inter-prefrontal suture usually weakly marked, but moderately marked in two specimens (MZUF 783, NMW 15301/2) (N = 13); supralabials 7 (3rd and 4th enter orbit) on either side of head (N = 17), but in BM 1902.12.13.85—although the head is damaged—it is evident that only the 2nd and 3rd supralabials on either side enter the orbit; anterior temporals usually two on either side of head (94.4%), but 1 left and 2 right in NMZB 17099 (N = 18); posterior temporals three on either side of head (38.5%), 3:4 or 4:3 (23.1%), 2:3 or 3:2 (23.1%), 1:2 in BM 1902.12.13.86 and 4 on right (left side damaged) in BM 1902.12.13.85 (N = 13); temporal formula on either side of head usually 2+3, occasionally 2+4 or 2+2; midbody scale rows 21–25 (mean 23.1 ±1.27, N = 19); ventrals 226–247 in males (N = 6), 241–271 in females (N = 10); subcaudals 57–68 in males (N = 4), 49–65 in females (N = 9). Dorsally brown D. abyssina have higher ventral + subcaudal counts than plain phase D. atra in Ethiopia (both sexes, 294–323 [N =7] versus 269–291 [N = 5] respectively). Ratio SVL: tail length 5.3–6.1 in males (N = 4), 5.5–8.1 in females (N = 10); total length: tail length 6.3–7.1 in males (N = 4), 6.5–9.1 in females (N = 10).
Size. Largest male (NMW 15301–3 – Eritrea) has a SVL of 504 mm (tail trunctated); largest female (MNHN 6567 – ‘Abyssina’, proposed lectotype, see below) 665 + 90 = 755 mm (see Duméril et al. 1854).
Habitat. Open savannah in the Ethiopian Highlands at elevations of 1900–2450 m (Figs 21 & 22). A subadult male, BM 69.11.4.38, although having an ‘Arabia’ (e.g. Gans 1959, Appendix 1) label in its museum jar, was in fact collected at Senafe, Eritrea at 2450 m (Blanford 1870), probably in Juniperus coniferous forest. Keriyo, north of Addis Ababa, is the southern-most locality for this species, situated at an altitude of 2320 m (Spawls 2004).
INSERT FIGS 21 & 22
Distribution. Restricted to central and north-western Ethiopia, and central Eritrea (Fig. 3).
Localities. (Plain phase specimens indicated by asterisks.) ERITREA. No locality, NMW 15301–1, 15301–2, 15301–3; Gheleb [NE 1538D4] NMW 15300; Senafe [NE 1439C2] BM 69.11.4.38. ETHIOPIA. No locality, MNHN 6567 (proposed lectotype, see below); Gondar [NE 1237C2] FMNH 12728, MSNM 2699a*,b,c, MZUF 783; Gondar to Asmara [ca. NE1338A = Ras Daschen Terera] MCZ 80984*; Keriyo [NE 0938B3] NMBO 10256–7*, NMZB 17099*; Mai Canetta [NE 1338D4] MSNM 2700a*,b*; Zegie, Lake Tsana [NE 1137C2] BM 1902.12.13.85–6*.
Biogeography. In the terrapin Pelomedusa subrufa Bonnaterre species complex, P. barbata Petzold et al. occurs in Saudi Arabia and Yemen, with closely-related populations of P. gehafie Rüppell in Eritrea and eastern Sudan, and P. somalica Petzold et al. in northern Somalia (Petzold et al. 2014). Similarly, D. arabica sp. nov. inhabits Saudi Arabia and Yemen, D. crucifera sp. nov. is found in northern Eritrea, D. abyssina is restricted to central Eritrea and the northern half of Ethiopia, while D. taylori sp. nov. occurs only in Somaliland and adjacent Djibouti.
Conservation. In terms of the regional (study area) and global distribution of this species, using IUCN (2012, 2016) criteria, it appears to be widespread and should be regarded as Least Concern.
Remarks. Saleh & Sarhan’s (2016: 26) comment that Trape et al. (2012) concluded that D. abyssina is a valid species “based on molecular phylogenetic data” is incorrect. Trape et al. (2012) did not include D. abyssina in their genetic analysis but followed Trape & Mané (2006) who treated this species as valid on morphological grounds.
Dasypeltis crucifera Bates sp. nov.
Cross-marked Egg-eater
Figs 23A–B, 24A–C & 25A–B
Dasypeltis scabra (not Linnaeus) Gans, 1959: 141 (part, Bogos, Pl. V, Fig. 4; Appendix 1, p. 204); Largen & Rasmussen, 1993: 340–341 (part, Agordat, Keren).
Holotype. ZMB 7631 (at Museum für Naturkunde, Berlin), an adult female (right side ovary examined) from Bogos, Eritrea (Gans 1959, Pl. V, Fig. 4; Fig. 23A–B & 24A–C). The specimen was purchased from Edward Gerrard (a London taxidermist and dealer of natural history objects) by the Zoologisches Museum der Königlichen Friedrich-Wilhelms-Universität zu Berlin during the second half of the 19th century (F. Tillack in litt., 15 April 2016). Bogos is a name that may refer to the city of Keren (also known as Senhit or Blin: 15°46’40”N, 38°27’29”E; NE 1538C4), but it may have been used as a more general term referring to the land of the Bogo people (an indigenous ethnic group) in Anseba Region, between the highlands and lowlands of Eritrea.
Paratypes. Two specimens: ERITREA. Allotype. MZUT 3582 (at Museo Regionale di Scienze Naturali di Torino, Turin), a juvenile male from Agordat [15°33’N, 37°53’E; NE 1537D2] (the specimen tag notes ‘Giachetti! cei det. 21.8.88’ [Fig. 25A–B]); MSNG 6738, probably female, from Keren [15°46’40”N, 38°27’29”E; NE 1538C4].
Additional material. One specimen: ERITREA. MZUT 3469, an adult female from Agordat [as above].
Diagnosis. Where data for MZUT 3469 (see “Additional material”) falls outside the ranges recorded for the type series, this is indicated below.
Assigned to the genus Dasypeltis on account of its slender form, possession of (usually 3–4) rows of reduced, oblique, keeled and serrated lateral scales (little or no serration in D. inornata) and head barely distinct from the neck (moderately distinct in D. fasciata). Differs from all other taxa in the genus by its unique dorsal colour pattern, in which dark brown oval saddles and lateral bars almost completely enclose pale cross-markings that form the inter-saddle areas (a variation of the ‘5I’ pattern found in Arabian Dasypeltis; Figs 23A & 25A), together with a combination of the following characters: Pattern cycles 51–62 (higher than D. bazi 38–49; lower or mostly lower than D. fasciata 66-141, D. abyssina 65-98, D. atra 62-130, D. arabica sp. nov. 62-79; D. inornatus always uniform brown above, D. gansi uniform or weakly marked); flanks decorated with dark vertical bars (spots or blotches in D. bazi, D. loveridgei comb. nov. and some populations of D. scabra); and venter immaculate apart from a few dark markings at the edges (extensive dark markings in D. abyssina and in many D. scabra); Ventrals: 226 in male (higher than D. taylori sp. nov. 196–216, D. sahelensis 207–221, D. inornata 208–225, D. parascabra 210–223), 231–243, 247 (MZUT 3469) in females (higher or mostly higher than D. taylori sp. nov. 208–222); Subcaudals 61 in male (higher than D. loveridgei comb. nov. 47–57; lower than D. inornata 81–92, D. fasciata 59, 71–91, D. medici 69–109, D. gansi 68–83, D. palmarum 68–77, D. parascabra 67–74, D. latericia 66–86, D. arabica 63–65), 49–52 in females (lower than D. inornata 69–84, D. fasciata 64–84, D. parascabra 57–64, D. palmarum 62–77, D. medici 61–90, D. gansi 59–73; D. latericia 59–72, D. arabica sp. nov. 53–61); Inter-prefrontal sulcus weakly marked (moderately to deeply marked in D. arabica sp. nov., weakly to moderately marked in D. bazi, usually moderately marked in D. taylori sp. nov., and variable in D. scabra [often well marked in the south of its range]); Frontal shield marginally pitted, State 1 or 2 (extensively pitted, States 3 & 4, in D. medici, D. taylori sp. nov. and many populations of D. scabra especially in the south of its range); Nasal divided below the nostril (undivided in D. sahelensis and D. parascabra); Postoculars 2 on either side of head (often one, especially in western populations, of D. atra); Supralabials usually 7 (3rd and 4th enter orbit) on either side of head (usually 6[2,3] in D. loveridgei comb. nov.).
Description of holotype. The holotype (ZMB 7631) is an adult female with a total length of 506 mm, snout-vent length 442 mm, tail length 64 mm. Ratio SVL: tail length 6.9, total length: tail length 7.9.
Head small and slightly distinct from the neck; snout rounded. Elongate nasal semi-divided (divided below the nostril), nostril pierced in anterior half of nasal, no loreal. Paired prefrontals larger than the paired internasals (individually and in combination). Inter-prefrontal sulcus weakly marked. Frontal large, widest anteriorly, slightly longer than broad, and smooth except for a single row of pits on the lateral borders (State 1). There is a single preocular and two postoculars on either side of head. Temporals 2+3 on either side of head. Seven supralabials, 3rd and 4th bordering the eye, on either side of head. First pair of infralabials in contact behind the mental, followed by a pair of large and elongate sublinguals in broad contact, and another pair of smaller sublinguals in narrow contact with one another and in contact with the first ventral plate, but separated posteriorly and medially by an oval extranumerary scale in broad contact with the first ventral plate. Eyes large (with vertically elliptical pupils), their greatest diameter much less than the distance from tip of snout to anterior part of eye socket.
Dorsal scales strongly keeled, unicarinate, in 25 rows at midbody, the third, fourth and fifth rows denticulate and obliquely placed. Dorsal scales in the anal region denticulate (see Gans 1959: 204). Vertebral scale row not enlarged. There are 231 ventrals, the anal plate is entire and there are 52 pairs of smooth subcaudals. After preservation in ethanol the dorsal colouration is pale brown with 62 ocelli or pattern cycles (markings are darker brown, saddles with paler brown centres) between nape and base of tail. Pale markings on the back are in the form of crosses, largely surrounded by brown, oval or diamond-shaped (sharpest ends anteriorly and posteriorly) dorsal saddles and similarly dark lateral bars, but with the tips of the crosses often separated from the dark markings. The tail has faint dark vertebral blotches anteriorly. Head with a chevron marking, interrupted on the left, with a dark arrowhead marking present on the nape. Venter immaculate apart from a few small brown spots extending from the outer scale row onto the outer edge of a ventral.
INSERT FIGS 23 & 24
Variation in paratypes (including allotype, as indicated). The allotype (MZUT 3582) is a juvenile male (211 + 39 = 250 mm; ratio SVL: tail length 5.4, total length: tail length 6.4); the paratype (MSNG 6738) is a female (368 + 45 = 413 mm, ratios 8.2 and 9.2). Both specimens have similar scalation and general appearance to the holotype, but differ as follows: frontal distinctly longer than broad in MSNG 6738; on the right side of the head of the allotype there are 2+2 temporals; pair of small posterior sublinguals in contact with first ventral plate and not separated by an extranumerary scale (also allotype); longitudinal rows of dorsals at midbody 22 in allotype, 24 in MSNG 6738; ventrals 226 in allotype (male), 243 in MSNG 6738 (female); pairs of subcaudals 61 in allotype (male), 49 in MSNG 6738 (female); pattern cycles on the back 52 in allotype, 51 in MSNG 6738; in allotype, forward-pointing chevron marking at the back of the head is followed by a dark oval marking that extends onto the nape, where it is followed by a backwards-facing chevron; in MSNG 6738 there is a forward-pointing chevron on top of the head, followed by a forward-pointing arrowhead on the nape with a connecting stalk extending onto the neck. The dark (brown) markings (often diamond-shaped) of the allotype are less distinct than in the holotype, and only barely distinguishable from the ground colour on the posterior part of the body and on the tail; in MSNG 6738 the dark oval saddles are often more rounded at the ends such that the pale crosses are less X-like in appearance. Serration of lateral scale rows and scales above the anal region, and condition of the anal plate, not determined. No live specimens are known to the authors, nor are any photographs available.
INSERT FIG. 25
Description of additional material: MZUT 3469 (female). This specimen is not currently available for examination and no photograph of it is available to determine its dorsal pattern (F. Andreone pers comm., 3 April 2016). However, it has 247 ventrals, 50 subcaudals and 23 midbody scale rows.
Size. Largest male (allotype, MZUT 3582 – Agordat) 211 + 39 = 250 mm; largest female (holotype, ZMB 7631 – Bogos) 442 + 64 = 506 mm.
Etymology. The name of the new species refers to the pale cross-marks between the saddles on its back.
Habitat. This species occurs in the hilly area between the lowlands associated with the drainage of the Barka River, the Sahel and the Highlands of Hamasien. Agordat is situated at low altitudes (about 600 m), while the city of Keren is at an altitude of about 1417 m.
INSERT FIG. 26
Distribution. Known from only two areas (Bogos/Keren and Agordat) in northern Eritrea (Fig. 3). Keren and Agordat are about 75 km apart.
Mimicry. Dasypeltis crucifera sp. nov. may mimic Echis megalocephalus Cherlin, 1990 (see photograph of “E. cf. megalocephalus” from Ginda in Dobiey & Vogel 2007; Fig. 30).
Biogeography. In the terrapin Pelomedusa subrufa Bonnaterre species complex, P. barbata Petzold et al. occurs in Saudi Arabia and Yemen, with closely-related populations of P. gehafie Rüppell in Eritrea and eastern Sudan, and P. somalica Petzold et al. in northern Somalia (Petzold et al. 2014). Similarly, D. arabica sp. nov. inhabits Saudi Arabia and Yemen, D. crucifera sp. nov. is found in northern Eritrea, D. abyssina is restricted to central Eritrea and the northern half of Ethiopia, while D. taylori sp. nov. occurs only in Somaliland and adjacent Djibouti.
Conservation. Using IUCN (2012, 2016) criteria and considering that it is known from only two areas, this species should be regarded as Data Deficient.
Remarks. The holotype (ZMB 7631) was listed under D. scabra (‘5I’ dorsal pattern) by Gans (1959) who failed to mention its distinctive cross-mark pattern. He also indicated that while other African specimens with the ‘5I’ dorsal pattern had heavily-marked venters (here referred to D. abyssina), this specimen had ‘immaculate’ underparts.
Dasypeltis arabica Broadley & Bates sp. nov.
Arabian Egg-eater
Figs 27A–C, 28A–B & 29A–B
Dasypeltis scabra (not Linnaeus) Boulenger, 1905: 180 (El Kubar, Yemen); Leviton & Aldrich, 1954, xxiii (checklist); Gans 1959: 141 (part, El Kubar) & 1961: 73 (part, Arabia); ? Corkhill & Cochran, 1965: 485 (Little Aden oil refinery); Spanò, 1972: 132 (El Siyani, Yemen); Gasperetti, 1988: 233, 410, Fig. 39 (1) (Ajibah, Saudi Arabia); Schätti & Desvoignes, 1999: 82; Schätti & Gasperetti, 1994: 386 (Aqabat Raydah & Abha, Saudi Arabia).
Dasypeltis saeizadi Hoser, 2013: 60 (“Saudi Arabia”) (nomen dubium); Göthel, 2015a: 19 & 2015h: 92; Saleh & Sarhan, 2016: 36.
Dasypeltis cf. scabra Sindaco et al. 2013: 93 (part, “SW Arabia and Yemen”).
Holotype. BM 1987.2192 (at Natural History Museum, London), an adult female (oviducts examined) from Sana’a, North Yemen [15º19’N, 44º14’E] collected by M. Al-Safadi (Figs 27 & 28).
Paratypes. Six specimens: YEMEN. Allotype. BM 1903.6.26.41 (at Natural History Museum, London; wrongly listed as ‘xBM 03-6-21-41’ by Gans 1959), an adult male (Gans 1959) from El Kubar, South Yemen [14º25’N, 45º01’E] collected by G.W. Bury; BM 1987.2193, adult female from Taiz, North Yemen [13º35’N, 44º01’E] collected by M. Al-Safadi; MSNG 52221, male from El Siyani, North Yemen, collected by G. Scortecci, 13 August 1965. SAUDI ARABIA. BM 1987.1018, subadult female from Ajibah [19º41’N, 41º57’E] collected by J. Gasperetti (Fig. 29); MHNG 2536.41, male from Aqabat Raydah [NE 1942C]; MHNG 2536.42, female from vicinity of Abha [about 18º13’N, 42º30’S].
Diagnosis. Assigned to the genus Dasypeltis on account of its slender form, possession of (usually 3–4) rows of reduced, oblique, keeled and serrated lateral scales (little or no serration in D. inornata) and head barely distinct from the neck (moderately distinct in D. fasciata). Distinguished from other congeners by its unique dorsal colour pattern in which black rectangular saddles and lateral bars form complete closure around the pale inter-saddle areas (Figs 27A & 29A; this pattern also occurs on parts of the back of a few D. abyssina from Eritrea, see pattern‘5I’ of Gans 1959, Pl. VI, Fig. 4); together with a combination of the following characters: Pattern cycles 62–79 (≤62 in D. crucifera sp. nov., D. taylori sp. nov. and D.bazi; D. inornatus always uniform brown above, D. gansi uniform or weakly marked); flanks decorated with dark vertical bars (spots or blotches in D. bazi, D. loveridgei comb. nov., and some populations of D. scabra—e.g. Fig. 5G); and venter immaculate apart from a few dark markings at the edges (extensive dark markings in D. abyssina and many D. scabra); High numbers of ventrals: 236–244 in males (higher than D. scabra 180–226, D. taylori sp. nov. 196–216, D. confusa 199–227, D. atra 199–232, D. sahelensis 207–221, D. inornata 208–225, D. bazi 213–229, D. parascabra 210–223 and D. crucifera sp. nov. 226), 239–254 in females (higher than D. taylori sp. nov. 208–222, D. sahelensis 212–237, D. bazi 215–232, D. inornata 219-237, D. parascabra 224–237); Subcaudals 63–65 in males (higher than D. loveridgei comb. nov. 47–57, D. bazi 59–61, D. crucifera sp. nov. 61; lower than D. inornata 81–92, D. fasciata 59, 71–91, D. medici 69–109, D. gansi 68–83, D. palmarum 68–77, D. parascabra 67–74, D. latericia 66–86), 53–61 in females (higher than D. loveridgei comb. nov. 39–50, D. taylori sp. nov. 44–52, D. crucifera sp. nov. 49–52; lower than D. inornata 69–84, D. fasciata 64–84, D. palmarum 62–77; mostly lower than D. medici 61–90); Inter-prefrontal sulcus usually moderately, occasionally deeply, marked (usually weakly marked in other species, but weakly to moderately marked in D. bazi, usually moderately marked in D. taylori sp. nov., and variable in D. scabra [often well marked in the south of its range]); Frontal shield marginally pitted, usually State 1 (extensively pitted, States 3 & 4, in D. medici, D. taylori sp. nov. and many populations of D. scabra especially in the south of its range); Nasal divided below the nostril (undivided in D. sahelensis and D. parascabra); Postoculars 2 on either side of head (often one, especially in western populations, of D. atra); Supralabials 7 (3rd and 4th enter orbit) on either side of head (usually 6[2,3] in D. loveridgei comb. nov.).
Description of holotype. The holotype (BM 1987.2192) is an adult female with a total length of 714 mm, snout-vent length 625 mm, tail length 89 mm; ratio SVL: tail length 7.0, total length: tail length 8.0.
Head small and barely distinct from the neck; snout rounded. Elongate nasal semi-divided (divided below the nostril), no loreal. Paired prefrontals slightly larger than the paired internasals (individually and in combination). Inter-prefrontal sulcus moderately marked. Frontal large, widest anteriorly, 1.14 times longer than broad, and smooth except for a single row of pits on the lateral borders (State 1). There is a single preocular and two postoculars on either side of the head. Temporals 3+3 (left), 2+4 (right). Seven supralabials, 3rd and 4th bordering the eye, on either side of the head. First pair of infralabials in contact behind the mental, followed by a pair of large and elongate sublinguals in broad contact which extend backwards to separate a smaller pair of posterior ones, which are followed by a pair of supranumerary shields, the right one being partially fused with the first ventral. Eyes moderate in size (with vertically elliptical pupils), their length much less than the distance from tip of snout to anterior part of eye socket.
Dorsal scales strongly keeled, unicarinate, in 25 rows at midbody, the third, fourth, fifth and sixth rows denticulate and (except for the 3rd row) obliquely placed. Dorsal scales in the anal region very feebly denticulate. Vertebral scale row not enlarged. There are 240 smooth ventrals, the anal plate is entire and there are 53 pairs of smooth subcaudals. After preservation in ethanol the dorsal colour pattern consists of black rectangular saddles and similarly coloured lateral bars (on a pale brown to grey background) that form complete closure around the pale (whitish) square to rectangular inter-saddle areas. There are 67 ocelli or pattern cycles (dark with pale centres) between nape and base of tail. The tail has faint dark vertebral blotches anteriorly. Head with remnants of fragmented chevrons, and a dark forward-pointing arrowhead marking is present on the nape. Venter immaculate cream to white apart from a few dark spots extending from the outer scale rows onto the outer edges of ventrals.
INSERT FIGS 27 & 28
Variation in paratypes (including allotype, as indicated). The six paratypes comprise three males and three females. The largest male (MSNG 52221) measures 563 (486 + 77) mm; BM 03.6.21.41 (allotype) measures 325 + 56 mm. Measured females (BM 1987.2193: 400 + 64 mm, BM 1987.1018: 246 + 60 mm) are smaller than the holotype. The paratypes have similar scalation and general appearance to the holotype, but differ as follows: Midbody scale rows are 23 (3 specimens, including allotype), 24 (1) or 25 (2) (Appendix 1). Ventrals 236–244 in males (244 in allotype), 239–254 in females (Appendix 1). Subcaudals 63–65 in males (65 in allotype), 53–61 in females (Appendix 1). Paired prefrontals much larger than the paired internasals in MHNG 2536.41 & 42; inter-prefrontal sulcus usually moderately developed but deeply marked in MHNG 2536.41; frontal with two rows of pits (State 2) on its outer surface in MHNG 2536.41; temporals in 2+3 arrangement in MHNG 2536.41 & 42; usually third, fourth and fifth (also sixth in two specimens) lateral scale rows reduced in size, serrated and inclined; serration on scales above vent faint or absent. Pupils appear round in BM 1987.2193. In MHNG 2536.41 the dark dorsal markings are joined along only the last one-fifth of the body (from snout to vent), elsewhere being similar to the ‘5N’ markings of D. scabra (see Gans 1959), while in MHNG 2536.42 most of the markings on the body are D. scabra-like, but with several ‘5I’ markings posteriorly. Dorsal pattern cycles 62–79 (Appendix 1), the first neck band divided in BM 03.6.21.41 (allotype) and MHNG 2536.42 (divided by an elongated vertebral band). A narrow, somewhat irregular, chevron is present on the back of the head in BM 1987.1018. No live specimens are known to the authors, nor are any photographs available. (Frontal and eye-snout measurements, and patterning on the top of the head, not recorded for all paratypes and additional material. For MSNG 52221 only size [see above], colour pattern, and numbers of supralabials, preoculars, postoculars, temporals, midbody scale rows, ventrals and subcaudals were recorded.)
INSERT FIG. 29
Size. Largest male (MSNG 52221 – El Siyani, North Yemen) 486 + 77 = 563 mm; largest female (BM 1987.2192, holotype – Sana’a, North Yemen) 625 + 89 = 714 mm.
Etymology. The name of the new species refers to the region of origin (Arabia) of the type series.
Habitat. This species occurs in the south-western highlands of Saudi Arabia and Yemen from 1000 m to 2100 m (annual rainfall exceeds 300 mm).
Distribution. South-western part of Saudi Arabia and western half of Yemen (Fig. 3).
Mimicry. Dasypeltis arabica sp. nov. may mimic Echis coloratus Günther (see photographs from the Sinai Peninsula in Dobiey & Vogel [2007]). Gasperetti (1988) showed that E. coloratus also occurred from Saudia Arabia to the south coast of Yemen, and illustrated a topotype from Jabel Grayyah in Saudia Arabia (his Fig. 123) showing pale vertebral spots (described as ‘pinkish’ in the description of the holotype by Günther [1878]). The new species is also similar to Echis megalocephalus (Fig. 30).
INSERT FIG. 30
Biogeography. Dasypeltis arabica sp. nov. has a similar distribution to Naja arabica Scortecci, except that the latter also occurs further east in the highlands of Dhofor, Oman (Trape et al. 2009). The geological history of the region is summarised by Leviton (1986): “During the entire Mesozoic and early Cenozoic, Arabia, Nubia and Somalia were assembled in one continental block at some times covered by shallow seaways (Bosellini, 1986). The onset of Neogene rifting, accompanied by the ‘slight oblique drifting’ of Africa and Arabia resulted in the opening of the Gulf of Aden.” Boulenger (1905) described, from El Kubar in Yemen, Atractaspis andersoni which was considered closely related to A. leucomelas Boulenger of Somalia, and also Zamenis [= Platyceps] variabilis, closely related to Platyceps brevis Boulenger of Somalia. In the terrapin Pelomedusa subrufa Bonnaterre species complex, P. barbata Petzold et al. occurs in Saudi Arabia and Yemen, with closely-related populations of P. gehafie Rüppell in Eritrea and eastern Sudan, and P. somalica Petzold et al. in northern Somalia (Petzold et al. 2014). Similarly, D. arabica sp. nov. inhabits Saudi Arabia and Yemen, D. crucifera is limited to northern Eritrea, D. abyssina is restricted to central Eritrea and the northern half of Ethiopia, while D. taylori sp. nov. occurs only in Somaliland and adjacent Djibouti. Lavin & Papenpuss (2012) cite several recent molecular studies that show that the Red Sea and Gulf of Aden are barriers to various reptiles between the Arabian Peninsula and Africa.
Conservation. Using IUCN (2012, 2016) criteria this species appears to be widespread and should be regarded as Least Concern.
Remarks. Corkill & Cochran (1965) recorded a specimen (whereabouts unknown, not plotted on map) of “D. scabra” from Little Aden (oil terminal) on the coast in southern Yemen, but its identity remains problematic.
We concur with the opinions, recommendations and proposals of Kaiser et al. (2013) and Kaiser (2014) regarding best taxonomic practices, and therefore reject the unscientific and unethical taxonomy of Hoser (2013, description of Dasypeltis saeizadi). We note specifically that the privately published and personally edited work of Hoser (2013) is in direct violation of the spirit and intention of the Code (ICZN 1999) as indicated by, inter alia, Recommendation 8A which explicitly encourages publication in “appropriate scientific journals or well-known monographic series” (see also comments by Schleip [2014] regarding non-compliance with the ICZN [1999]). It can be added that in their Snakes of the World catalogue, Wallach et al. (2014: ix) state: “The Australasian Journal of Herpetology [in which the Hoser 2013 paper was published], of which Hoser is the editor, reviewer, publisher, and sole author, is not considered a valid publication under the spirit of the Code and therefore all proposed names are nomina illegitima (see Wallach et al., 2009 and Kaiser et al., 2013).”
Hoser (2013) named ‘BMNH – 1987.1018’ as holotype and sole type specimen of what he considered a new species, namely D. saeizadi, from ‘Saudi Arabia’. His diagnosis is based entirely on a vague comparison of colour patterns be tween D. scabra, D. abyssina (presumably based on MNHN 6567, the Ethiopian syntype of R. abyssinus – the only specimen of this taxon known at the time, incorrectly referred to as the ‘holotype’ of D. abyssina by Trape & Mané 2006, see discussion above) and the proposed new species. Hoser (2013: 60) noted that the latter is “readily separated” from both D. scabra and D. abyssina “by the dorsal colouration”. However, he proceeded to make the following erroneous statements in this regard: According to Hoser (2013: 60): D. abyssina has “indistinct markings on the head and body and a complete absence of barring on the upper labials, or if present, it [barring on supralabials] is effectively indistinct.” However, the proposed lectotype (MNHN 6567) of D. abyssina and all other known (patterned) specimens of this species (see species account above) have very distinct dark markings dorsally and laterally on the head, body and tail, including distinct barring on the supralabials (see Duméril et al. 1854, Pl. 81, Fig. 1 in colour & Fig. 20 in the current paper); while even the Nil Blanc ‘syntype’ (proposed paralectotype, MNHN 7227 = D. scabra) has distinct markings as described above. Hoser (2013: 60) then stated that his new species is “more similar to D. scabra in dorsal colouration, but lacks the distinctive whitish patches interspercing the dorsal midline blotches or squares. In turn these blotches or squares tend somewhat towards being pointed at the anterior end of the body”. Again this is incorrect as the dorsal pattern of BM 1987.1018 differs distinctly from D. scabra (Gans’ 1959 ‘5I’ versus ‘5N’ dorsal colour patterns respectively), the pale patches on its back are very distinct, and the saddles are for the most part not at all markedly pointed anteriorly (Fig. 29A). Hoser (2013: 60) added that his proposed new species “In contrast to the other two species [D. scabra and D. abyssina]” has a “darkening on the upper temples” and “Like D. abyssina there is a complete absence of barring on the upper labials”, “or if present, it [barring on the supralabials] is effectively indistinct.” Once again this is incorrect as Fig. 29A–B clearly shows that the upper temples of BM 1987.1018 are not darker than other parts of the head, nor are they noticeably darker than in D. abyssina (Duméril et al. 1854, Pl. 81, Fig. 1; Fig. 20C–G) or D. scabra (e.g. Gans 1959, Pl. IV-V; Broadley 1990, Fig. 152; Bates et al. 2014, p. 411, lower figure; Fig. 5), and the supralabials (of BM 1987.1018) are in fact distinctly barred, as are those of D. abyssina (Duméril et al. 1854, Pl. 81, Fig. 1; Fig. 20C–G). Hoser (2013) did not provide any additional features for distinguishing his proposed new species. It is therefore evident from the afore-mentioned that Hoser’s (2013) description is a mis-representation of reality—his description is not based on the true features of the specimen he names as holotype. His description cannot therefore be considered valid and in accordance with the requirements of the Code (e.g. Article 1, which requires that an actual specimen be used for the description of an extant or extinct species of animal). More specifically, Article 1.3.1 excludes from the provisions of the Code any name proposed for a hypothetical concept, which is what Hoser’s (2013) name must be considered, as it can be shown (see above) that it is not based on the features of the specimen he named as holotype (no paratypes).
According to P. Campbell at the Natural History Museum, London (in litt. 3 September 2015), no record exists of Raymond Hoser having ever borrowed any Dasypeltis from that institution, or of him having spent time at that museum examining such specimens. If Hoser had indeed examined the specimen, or even an image thereof, the characteristic ‘5I’ dorsal pattern (see Gans 1959, and Figs 27A & 29A) of Arabian populations of Dasypeltis would have been obvious immediately. Hoser (2013) also disregarded Recommendation 73C of the Code (1999) by not providing full collection details for the specimen and neglecting to provide any supporting meristic or morphometric data, including ventral, subcaudal and midbody scale row counts—all considered standard fare in modern snake descriptions. The lack of any such data in his description is a further indication that he did not examine the specimen (or an illustration thereof) that he named as holotype. In consideration of the above, Hoser’s (2013) name must be considered a species inquirendum and a nomen dubium and rejected as an available name for D. arabica sp. nov.
Multivariate analyses
For the analysis of all populations, two-way permanova (10 000 permutations) detected that there is no interaction between gender and species (df 9, F = –16.69, p = 0.67). However, it detected significant differentiation at species level (df 9, F = 2.43, p < 0.001) and at gender level (df 1, F = 5.28, p < 0.001) respectively, justifying separate analysis of sexes. For the Horn of Africa analysis, two-way permanova (10 000 permutations) detected that there is no interaction between gender and species (df 9, F = –3.19, p = 0.53). However, it detected significant differentiation at species level (df 6, F = 1.29, p < 0.001) and at gender level (df 1, F = 2.24, p < 0.001) respectively, justifying separate analysis of sexes.
A total of 249 specimens (D. medici: 9 males, 20 females; D. confusa: 6, 13; D. scabra: 24, 40; D. bazi: 3, 2; D. taylori: 3, 2; D. atra: 40, 62; D. fasciata: 2, 3; D. abyssina: 4, 7; D. crucifera: 1, 2; D. arabica: 2, 4) had complete sets of data and were used in the analyses (separately for males and females) of the entire study area. A total of 64 specimens (D. scabra: 6 males, 13 females; D. bazi: 3, 2; D. taylori: 3, 2; D. atra: 7, 9; D. abyssina: 4, 7; D. crucifera: 1, 2; D. arabica: 2, 4) with complete data were used in analyses (separately for sexes) of the Horn of Africa and Egypt. In the analyses for males from the latter area, numbers of preoculars was invariable and was therefore excluded. For pattern cycles, plain coloured specimens (including ‘intermediate’ D. medici with barely distinguishable darker dorsal markings) were scored as ‘0’.
Principal Component Analysis for north-eastern Africa and south-western Arabia
In the analysis of males (Fig. 31) the first two principal components (PC) explained most of the variance (36.2%). Only D. medici formed a distinct cluster distinguishable from all other populations, mostly along PC 2 for which numbers of subcaudals (0.54) and ventrals (0.40) had the highest loadings (Table 3). For females the first two components also explained most of the variance (37.2%), and again only D. medici formed a distinct cluster distinguishable from all other populations (Fig. 31), with the highest loadings (on PC 2) for subcaudals (–0.49) and pattern cycles (0.49) (Table 4). However, both sexes of D. scabra, D. atra and D. confusa also formed clusters and were partly distinguishable along PC 2.
INSERT FIG. 31
INSERT TABLES 3 & 4
Discriminant Function Analysis for north-eastern Africa and south-western Arabia
As all groups (as male or female of a species) are real geographical entities it is appropriate to conduct Discriminant Function Analysis to determine whether or not they are distinguishable. In the analysis of males of the various species (Fig. 32), 83.9% of specimens were correctly classified, but this included only 50% of D. confusa, while one out of three specimens of both D. taylori sp. nov. and D. bazi were mis-classified (Table 5). Dasypeltis medici was easily distinguishable from all other species along axis 1, on which numbers of subcaudals (0.81) and ventrals (0.60) load most heavily; Dasypeltis scabra and D. atra are largely separated along axis 2, on which fontal pitting (0.67) and pattern cycles (0.54) load most heavily; D. abyssina and D. arabica are also separated from D. scabra along axis 2, on which number of ventrals (0.41) is also an important factor (Table 6). For females (Fig. 32), 75.5% of specimens were correctly classified, but this included only 61.5% of D. confusa, 50% of D. taylori sp. nov., D. arabica sp. nov. and D. crucifera sp. nov., and neither of two specimens of D. bazi (Table 7). Dasypeltis medici is again easily distinguishable from all other species along axis 1, on which numbers of subcaudals (0.92) loads most heavily; Dasypeltis scabra and D. abyssina are well separated along axis 2, on which fontal pitting (0.67) and ventrals (0.64) load most heavily; and D. scabra and D. atra are partially separable on this axis (Table 8).
INSERT FIG. 32
INSERT TABLES 5–8
Principal Component Analysis for the Horn of Africa and Egypt
In the analysis of males the first two principal components (PC) explained most of the variance (40.1%); for females these components also explained most of the variance (38.7%). Although some species are largely distinguishable, none of the groups for males or females formed distinct clusters (Fig. 33; Tables 9 & 10).
INSERT FIG. 33
INSERT TABLES 9 & 10
Discriminant Function Analysis for the Horn of Africa and Egypt
In the analysis of males (Fig. 34) all specimens with the exception of one of three D. bazi were correctly classified, suggesting that they are 100% distinguishable in morphometric space (Table 11). All species were fully or partly (D. abyssina and D. arabica sp. nov.) distinguished along axis 1—on which ventrals (2.51) and midbody scale rows (2.09) load most heavily—except D. atra and D. bazi which are nevertheless largely separated along axis 2, for which pattern cycles (0.82) and frontal pitting (–0.68) load most heavily (Table 12). For females (Fig. 34), 81.6% of specimens were correctly classified; this figure was at least 75% for all species except D. bazi for which one of two specimens was incorrectly classified (Table 13). Dasypeltis scabra was distinguished from D. arabica sp. nov. and D. abyssina along axis 1, while the latter two species were distinguished along axis 2; D. taylori sp. nov. and D. bazi were distinguished from D. arabica sp. nov. and D. abyssina along axis 1; D. scabra is also distinguished from D. crucifera sp. nov. along axis 2; D. atra is distinguished from D. arabica sp. nov. along axis 2, and also partly from D. scabra on this axis (Table 14). Ventrals (1.07) load by far the most heavily on axis 1, whereas subcaudals (1.02) and pattern cycles (0.68) load most heavily on axis 2.
INSERT FIG. 34
INSERT TABLES 11–14
SUMMARY OF SPECIES
A morphological analysis of the genus Dasypeltis in north-eastern Africa and south-western Arabia indicates that as many as 10 species occur there. This includes three new species (D. taylori sp. nov., D.crucifer sp. nov. and D. arabica sp. nov.) previously subsumed under the name D. scabra, as well as two (D. abyssina and D. bazi) that had until recently also been considered referable to D. scabra. The ranges of these five species do not overlap. This taxonomy is partly supported by the multivariate analyses of scale characters and numbers of pattern cycles, and colour pattern is often diagnostic of species.
Dasypeltis medici (Bianconi)
Although it occurs as patterned, weakly patterned and unicolour morphs (dorsally), this species is easily distinguishable by the presence of paired apical pits—on the dorsal scales—which are sharply defined by dark pigment, an extensively pitted frontal shield, and high subcaudal counts (75–109 in males, 61–90 in females). Also, patterned specimens have 1–5 distinct, often narrow, chevrons on the neck (one or two similar chevrons may be present in some other species). In the study area this species is restricted to southern Somalia, south-eastern Kenya, and north-eastern Tanzania; found along the coast but also in some savannah and mountainous areas.
Dasypeltis confusa Trape & Mané
Although often similar in scalation characteristics to D. scabra and D. atra, with which it is sympatric and parapatric, this species is easily distinguished by its colour pattern of large diamond-shaped saddles (often outlined in white) confluent with the lateral bars. In the study area it occurs only in the west (southern South Sudan, Uganda, western Kenya, Rwanda).
Dasypeltis scabra (Linnaeus)
A variable species widely distributed in the study area, from Burundi and Tanzania northwards to southern Somalia and central Ethiopia, with isolated populations along the Nile River in South Sudan and Sudan. It always has the ‘5N’ dorsal pattern, and differs from patterned D. atra by seldom having transversely enlarged saddles on the back, and usually also by having a more boldly marked head. There are usually two postoculars on either side of the head, while D. atra often has only one.
Dasypeltis bazi Saleh & Sarhan
This species has a distinctive colour pattern consisting of large cranio-caudally elongate saddles, usually bifurcated at each end with a fairly shallow but distinct notch, and the flanks are decorated with spots or blotches (not bands or vertical bars). The only locality at which this species is currently known to occur, namely the Fayoum Depression, is separated from other populations of Dasypeltis by over 1 300 km.
Dasypeltis taylori Bates & Broadley sp. nov.
This species, characterised by a combination of features including a strongly pitted frontal scale, is found in Somaliland and Djibouti, separated from populations of D. scabra to the south by the arid Ogaden plateau. The dorsal saddles are often large, closely-spaced, square or slightly rectangular, and separated by narrow, usually sharply defined, white crossbars; there are low numbers of pattern cycles (44–59).
Dasypeltis atra Sternfeld
Occurs as three colour phases: brown dorsally and cream to yellow or orange ventrally; black dorsally (unique in the genus) and grey ventrally; and patterned, sometimes similar to the ‘5N’ pattern of D. scabra but more typically with transversely widened saddles, often in contact with the lateral bars. Typically the frontal scale is only marginally pitted and the inter-prefrontal suture weakly marked; and western populations usually have only a single postocular (rather than two), a condition rare in others of this genus. This species is widespread in the study area, from northern Tanzania to Burundi, Rwanda, Kenya, Uganda, south Sudan and Ethiopia as far north as Circao. In some areas it is sympatric and/or parapatric with D. scabra, D. confusa, D. medici, D. fasciata and D. abyssina.
Dasypeltis fasciata A. Smith
Dorsum patterned, with body pattern always expressed as chequerboard on skin between scales and often also on lower edges of these, rather than on most of the scales themselves as in patterned specimens of other species. Also characterised by high subcaudal counts (59, 71–76 in males, 64–76 in females). In the study area this highly arboreal species is restricted to forests in western Uganda.
Dasypeltis abyssina (Duméril, Bibron & Duméril)
Easily recognisable by the combination of its unique dorsal pattern (including oblique transversely widened saddles which may merge to form zigzags on the back) and heavily marked venter. This species also has the highest ventral counts in the genus (241–271 in females, 226–247 in males). It is restricted to the Ethiopian highlands that extend into Eritrea.
Dasypeltis crucifera Bates sp. nov.
Found near the northern limits of D. abyssina, but at lower altitudes in northern Eritrea below the Ethiopian highlands. Distinguished by its unique dorsal pattern in which dark brown oval saddles and lateral bars almost completely enclose pale cross-markings that form the inter-saddle areas; venter largely unmarked (D. abyssina, its nearest congener, has a markedly different dorsal pattern and a heavily marked venter).
Dasypeltis arabica Broadley & Bates sp. nov.
Easily distinguished by its unique dorsal pattern consisting of black rectangular saddles and lateral bars that form complete closure around the pale inter-saddle areas (pattern ‘5I’ of Gans 1959); the venter is mostly unmarked. It has high ventral counts. This is the only species in the genus restricted to Arabia, where it occurs in the highlands of Saudi Arabia and Yemen.
DISCUSSION
Dasypeltis scabra is by far the most widespread species in the genus, and the most widely distributed of the 10 species found in north-eastern Africa and south-western Arabia. It ranges from the Cape of Good Hope through large parts of southern, central (excluding forests) and eastern Africa (‘5N’ dorsal pattern of Gans 1959; Broadley 1990; this study), with apparently isolated populations along the Nile River (Sudan and South Sudan) and near Kutum (western Sudan). The isolated Egyptian population in the Fayoum Depression is now considered a separate species (D. bazi), as are populations in Somaliland/Djibouti (D. taylori sp. nov.), northern Eritrea (D. crucifera sp. nov. ), the northern Ethiopian highlands (D. abyssina) and western Arabia (D. arabica sp. nov.). Dasypeltis confusa has a large range in western and central Africa (‘5L’ of Gans 1959), extending as far east as South Sudan, Uganda, western Kenya and Rwanda. Dasypeltis fasciata is arboreal and restricted to forests but has a large range in western and central Africa, extending as far east as Uganda. Dasypeltis medici, now considered a monotypic species, occurs in coastal and inland forests from southern Somalia to northern KwaZulu-Natal in South Africa; it also occurs in savannah where it may be sympatric with D. scabra. Dasypeltis atra, at one time considered a montane forest subspecies of D. scabra, is clearly a separate species (e.g. Gans 1964) with an extensive range from eastern Democratic Republic of the Congo to Rwanda, Burundi, Uganda, South Sudan, Kenya and the Ethiopian highlands. The range of D. atra is overlapped almost entirely by that of D. scabra, and although the two species appear to be mostly parapatric, the former also occurs in some savannah areas (e.g. Broadley & Bates 2009) where it is sympatric with the latter.
Gans (1959) considered that evolutionary relationships might be indicated by how dorsal colour patterns were derived. For example, he demonstrated how a generalised ‘5N’ D. scabra pattern could have given rise to the ‘5L’ D. confusa pattern, leading to the D. palmarum pattern (saddles and lateral bars confluent and much narrowed with increased inter-saddle areas), and how the marked D. medici pattern could have been derived from ‘5N’. His diagrammatic scheme of evolutionary history, in which the more derived species are smaller (body length), have better developed lateral body serration, and increased pitting on the head, places D. inornata as the basal species, followed by D. fasciata, “montane” (= D. atra), D. palmarum, and ‘D. scabra’ (including ‘5N’ = D. scabra, ‘5L’ = D. confusa and ‘5I’ = D. arabica sp. nov.), with D. medici as the most derived species. Gans (1959: 172) noted that D. scabra may have been derived “separately from the montane population [= D. atra], representing an invasion of the savannah habitat from the forest, and D. palmarum would be derived from scabra as a form adapted to the coastal dry belt of central West Africa.” However, Gans (1959: 173) warned that in Dasypeltis “…the use of different characters permitted the erection of sharply differing phylogenies.”
It is now generally accepted that the genus Dasypeltis, along with the mainly African genera Crotaphopeltis, Telescopus, Dipsadoboa, Toxycodryas and Boiga, is part of the Boigini clade of the colubrid subfamily Colubrinae (Gravlund 2001; Kelly et al. 2003; Lawson et al. 2005; Figueroa et al. 2016). Relationships within the genus, however, are not well understood, with only three published molecular phylogenies, one (16S only) dealing mainly with West African species (Trape et al. 2012), another using the same data but with two samples of D. bazi from Egypt added (Saleh & Sarhan 2016), and a multigene study that used the same species as Trape et al. (2012) but added additional samples, including three ‘D. atra’ (Figueroa et al. 2016). The phylogeny of Trape et al. (2012) indicated that D. confusa (samples from Togo) was the sister group to all other West African species (including D. fasciata) as well as D. scabra from ‘South Africa” and ‘D. cf. lineolata’ (possibly D. atra) from Burundi. Trape et al.’s (2012) study supported species status for all West African taxa (including D. latericia, previously considered a subspecies of D. gansi). It also suggested a close relationship between D. fasciata, D. gansi and D. latericia, and between D. sahelensis and D. parascabra. With the inclusion of one additional species, D. bazi, Saleh & Sarhan’s (2016) phylogeny suggested different relationships, but most of these were not well supported (e.g. D. scabra was paired with D. latericia, and D. confusa paired with D. cf. lineolata [? D. atra]). The phylogeny of Figuero et al. (2016) identified two main clades, one with D. confusa + D. fasciata versus the rest, but D. fasciata was also included in the larger clade. This latter clade was divided into two well supported subclades, one consisting of D. atra, D. scabra and D. latericia, and the other containing D. atra, D. gansi, D. fasciata and D. sahelensis. The confounding positions of both D. fasciata and D. atra in different clades may suggest the presence of cryptic species, but alternatively it may reflect incorrect identification of specimens. The relationships indicated by these studies may change or be resolved further with greater taxon sampling, particularly of north-eastern, eastern and southern African lineages (A. Barlow et al. in prep.).
Geniez & Guillod (2003) considered the populations of Dasypeltis in Morocco, Egypt and south-western Arabia as being Afrotropical relicts. Dasypeltis bazi appears to have a relict distribution indicative of a former widespread range in northern Africa that was reduced and fragmented following dessication of the region during the Holocene (Kuper & Kröpelin 2006). Oases such as the Fayoum appear to be vestiges of what was once an expansive savannah, probably with riparian-fringed links like the Nile River, and may act as climatic refuges. The geographical ranges of some snakes in Egypt, such as Telescopus dhara obtusus (Reuss), are closely associated with the Nile Valley and this species also occurs in apparent isolation on the Red Sea coast in the south-east of the country. Its distribution range may be similar to the past distribution of D. bazi, a species that is perhaps also represented by an apparently isolated population near the Red Sea coast at Erkowit.
During the Last Glacial Maximum and terminal Pleistocene (21000–8500 B.C.E.) the Sahara desert extended about 400 km further south than it does now, probably incorporating, inter alia, all of modern-day Sudan (Kuper & Kröpelin 2006). Monsoon rains that started 8500 B.C.E. resulted in hyperarid conditions being replaced by savannah-like environments, similar to the current-day Sahel, with formation of lakes and temporary rivers (Baha El Din 2006), perhaps allowing Dasypeltis (and many other animal species) from areas in central Africa to disperse and establish populations as far north as Morocco and Egypt. However, retreating monsoonal rains from 5300 B.C.E. resulted in dessication of the Eastern Sahara, with a return to full desert conditions throughout Egypt by 3500 B.C.E. (Kuper & Kröpelin 2006). Most Afrotropical animal species (or at least most populations) would then have retreated southwards as the only favourable (non-desert) habitat remaining in the region would have been in the Nile Valley and at scattered oases (Baha El Din 2006) such as Fayoum, Siwa, Qara, Bahariya, Farafra, Dakhla and Kharga. Although populations of Dasypeltis in Egypt (Fayoum and possibly in the Nile Valley itself) could be relics of a former widespread species that became established there during favourable conditions, they may alternatively be the descendants of populations that dispersed north along the Nile Valley in later times, using the valley as a ‘green corridor’ (see also Escoriza 2010). According to Baha El Din (2006), the Nile River (and its associated valley) varied considerably in size and impact during the Quaternary, before achieving its current form; and its function as a conduit for African fauna and flora was established as recently as 800 000 BP. Saleh & Sarhan’s (2016) study indicated that divergence times between D. bazi and other African Dasypeltis species predate the formation of the Nile River by about a million years, and this together with the absence of Dasypeltis records in the Nile Valley [in Egypt], suggests that the occurrence of these snakes in Fayoum is the result of vicariant events related to the formation of the Sahara rather than by dispersion across this arid desert.
Geniez & Guillod (2003) were of the opinion that while the Nile Valley represented a northward corridor of the Ethiopian region for some species of sub-Saharan reptiles, the Moroccan population (now known as D. sahelensis, with most of its range in the Sahel; Trape & Mané 2006) is isolated. According to Escoriza (2010), the occurrence of ‘D. scabra’ and ‘Boaedon fuliginosus’ (Boie) in northern Africa may be the result of range expansions during humid paleoclimatic conditions in the late Pleistocene or recent Holocene. Escoriza (2014) suggested that populations of D. sahelensis crossed the Western Sahara desert from the Sahel (or associated savannahs) to northwestern Africa (Western Sahara and Morocco) in humid conditions 6000–9000 years ago during the Holocene. This species, together with populations of the widespread snakes ‘B. fuliginosus’ and Bitis arietans (Merrem), was subsequently isolated in the relatively moist Atlantic coastal region of north-western Africa during the next period of aridification (Schleich et al. 1996; Geniez & Guillod 2003).
The nearest record to D. bazi from the Fayoum Oasis in Egypt is Erkowit in eastern Sudan near the Red Sea coast, but the latter town is situated about 1330 km away to the south-east. The specimen from Erkowit (D. cf. bazi) may represent an isolated relict population of D. bazi. This specimen is similar to Egyptian specimens of D. bazi, both in terms of colour pattern and scalation. This Egyptian/Erkowit population may in the past have been part of a more widespread population of D. scabra that became isolated following aridification of the Sahara during the Holocene, resulting in the evolution of a new species, D. bazi. The two populations may then have become isolated as climatic and environmental conditions changed, with the Fayoum Oasis (D. bazi) and Red Sea Hills (D. cf. bazi = Erkowit population) functioning as refuges, and accounting for their large current-day separation. Erkowit is a small town situated on a minor plateau (mostly 1000–1200 m a.s.l.) in the Red Sea Hills—a northern extension of the Ethiopian Plateau—at the eastern edge of the arid Sahel. Sea mist is blown in by north-easterly winds during winter to provide a moisture-temperature gradient at Erkowit, which has evergreen Afromontane vegetation only 10 km from the surrounding desert shrub (Vetaas et al. 2012). The Erkowit area was once regarded as a mist oasis in the desert, representing a potential refugium for Dasypeltis. Baha El Din (2006) suggested that some species of Afrotropical reptiles extended their ranges into Saharan Africa using the Nile River (and its associated valley) and Red Sea coast and mountains as conduits, although some populations in these areas are merely relicts. The cobra Naja nubiae Wüster & Broadley, 2003, for example, has a patchy distribution in eastern Chad, Sudan, Eritrea and at sites along the Nile River in Egypt.
Corkhill (1935) noted that in Sudan, D. scabra occurred only in ‘Blue Nile Province’—a region which his map indicates as being situated slightly east and south of Khartoum—but he did not list any specific localities. There are in fact four known Dasypeltis records for Sudan and one for South Sudan (none appear to be referrable to D. bazi), all but one located at or near the banks of the White Nile River. The exception is the newly-reported specimen (ZFMK 32479) from 50 km north of Kutum in Sudan, situated 1750 km from Fayoum and 1395 km WSW of Erkowit, identifiable as D. scabra (see above). One of the two syntypes of Rachiodon abyssinus (= Dasypeltis abyssina), collected in the mid-19th century during an expedition to the sources of the White Nile under the direction of M. d’Arnaud, and labelled “Nil Blanc” (= White Nile, possibly above Khartoum in Sudan), was examined by the author and is in fact a D. scabra (MNHN 7227, female, ‘5N’ dorsal pattern – dark-bordered brown saddles with vertically-elongate lateral bars between them, 54 pattern cycles, 226 ventrals, 53 pairs of subcaudals; Fig. 6). Khartoum, where the White and Blue Niles converge, is about 600 km south-west of Erkowit. A specimen (BM 09.10.15.35) of D. scabra from ‘Sennar State’ (which straddles the White Nile; Fig. 3) in south-eastern Sudan has some ‘5N’ markings dorsally, but also the peculiar ‘5I’ dorsal pattern over parts of its back (photograph examined) as described by Gans (1959) for some populations of Eritrean and Arabian ‘D. scabra’, where the dark saddles and lateral bars are joined, enclosing a pale inter-saddle region. This female specimen was in fact examined by Gans (1959) who recorded 65 pattern cycles, 227 ventrals and 52 pairs of subcaudals. The fourth Sudanese specimen (RMNH 24399, female) was collected 40 km north of Rabak, near Kosti (Fig. 3), on the banks of the White Nile and has the typical ‘5N’ D. scabra dorsal pattern, 50 pattern cycles, 222 ventrals and 55 subcaudals. There is one additional D. scabra record (FMNH 134939: male, ‘5N’ pattern, 64 pattern cycles, 213 ventrals, 62 pairs of subcaudals) on the White Nile (east bank) at Melut in the north-eastern part of South Sudan. Other records of Dasypeltis in adjacent parts of Eritrea and north-western Ethiopia are referable to D. taylori sp. nov., D. abyssina, and D. atra (see below). The four above-mentioned ‘5N’ D. scabra records on the White Nile, possibly representing a single population, appear to be considerably isolated from other populations of this species. The northern-most D. scabra record in the Nile Valley is Sennar State, about 700 km south-west of Erkowit. Apart from Erkowit, the nearest other D. scabra locality is the Addis Abeba area (Largen & Spawls 2010; Fig. 3) about 740 km ESE of Melut. There is one additional (apparently isolated) record of D. scabra from Wadelai, much further upstream, on the Albert Nile in north-western Uganda (Fig. 3).
As discussed in the species account, some Eritrean specimens of D. abyssina have the ‘5I’ dorsal pattern typical of D. arabica sp. nov. over parts of their backs. This, together with other morphological similarities (e.g. high numbers of ventrals and subcaudals) between D. abyssina and D. arabica, together with their geographical proximity, suggests a close evolutionary relationship. A continuous population may have been separated by the opening of the Gulf of Aden (see Leviton 1986), with subsequent divergence of the resultant sub-populations. The dorsal colour pattern of D. crucifera sp. nov. is similar to this ‘5I’ pattern and the close geographical proximity suggests that it may be closely related to the high-elevation D. abyssina. Dasypeltis taylori sp. nov. is separated from the nearest populations of D. scabra—a species it most resembles—by the arid Ogaden plateau, and may represent a relictual population of a once widespread D. scabra.
A molecular phylogeny of the genus, including samples of all species and with reasonable coverage of their ranges, will go a long way towards elucidating evolutionary relationships. Considering the vast areas covered by the ranges of most species, the possibility of additional (possibly cryptic) species cannot be discounted. Sampling of all species may be difficult at this time as some parts of Africa are still relatively inaccessible, not only physically, but in some cases also as a result of armed conflicts (e.g. parts of Somalia, Sudan, South Sudan, Eritrea-Djibouti border). Relationships and levels of divergence for the various isolated populations of D. scabra (e.g. 50 km N of Kutum, Sudan; populations along the Nile River in Sudan/South Sudan; Wadelei, Uganda) and D. cf. bazi (Erkowit, eastern Sudan) should also be investigated.
Key to the genus Dasypeltis in north-eastern Africa and south-western Arabia
1a. Paired apical pits on dorsal scales sharply defined by dark pigment; frontal shield extensively (State 3) to entirely (State 4) pitted; patterned specimens with 1–5 distinct chevrons on neck; subcaudals 75–109 in males, 61–90 in females ………………… D. medici
1b. Paired apical pits on dorsal scales not sharply defined by dark pigment; frontal varies from smooth to extensively pitted (States 0–4); when present, chevrons restricted to nape ………………………………………………………………………………………………………………………….. 2
2a. Dorsum patterned, with body pattern always expressed as chequerboard on skin between scales and often also on lower edges of these; frontal usually marginally pitted (States 1–2); lateral scales always strongly serrated; subcaudals 71–76 (once 59) in males, 64–76 in females ………………………………………………………………………………………………… D. fasciata
2b. Dorsum without pattern or with pattern expressed across the scales; frontal varies from smooth to extensively pitted (States 0–4); lateral serration present in varying degrees; subcaudals 47–73 in males, 43–65 in females ………………………………………………………… 3
3a. Dorsum uniform black, brown, grey or red, or pale brown or grey with a pattern of dense, dark brown, olive or reddish, usually transversely-enlarged, saddles, often joined to the lateral bars; when present 62–130 (mean 92.6) pattern cycles from nape to base of tail; postoculars 1 or 2 on either side of head ……………………………………………………….. D. atra
3b. Dorsum light brown or grey, uniform, or with a pattern of darker dorsal markings linked or alternating with lateral bars; 39–98 pattern cycles from nape to base of tail; usually 2 postoculars on either side of head …………………………………………………………………………. 4
4a. Dark dorsal markings (saddles) usually large and diamond-shaped, linked on either side to the dark lateral bars (or dorsum uniform pale brown); ventrals 199–224 in males, 212–239 in females ………………………………………………………………………………………. D. confusa
4b. Dark dorsal markings (saddles) variable in size but not diamond-shaped, usually not linked on either side to the dark lateral bars (or dorsum uniform pale brown in D. abyssina, but then ventrals 226 in males, 241–263 in females) …………………………………. 5
5a. Dark dorsal markings (saddles) and lateral bars completely or largely enclose a pale saddle patch; venter unmarked except for small dark specks at the lateral edges; frontal marginally pitted (States 1 & 2). …………………………………………………………………………… 6
5b. Dark dorsal markings (saddles) and lateral bars mostly separated, not enclosing a pale saddle patch; markings on venter variable; frontal pitting variable…………………………….. 7
6a. Pale markings on the back more-or-less square and completely surrounded by dark rectangular dorsal saddles and lateral bars; pattern cycles on back 62–79; inter-prefrontal sulcus moderately to deeply marked; ventrals 236–244 in males, 239–254 in females; subcaudals 63–65 in males, 53–61 in females ………………………………. D. arabica sp. nov.
6b. Pale markings on the back in the form of crosses, largely surrounded by dark oval dorsal saddles and lateral bars, but with the tips of the crosses often separated from the dark markings; pattern cycles on back 51–62; inter-prefrontal sulcus faintly marked; ventrals 226 in males, 231–247 in females; subcaudals 61 in males, 49–52 in females ……………………………………………………………………………………………… D. crucifera sp. nov.
7a. Dorsal markings (saddles) often oblique and wider than long, almost confluent or forming zig-zags, usually not linked to lateral bars; parts of the labials and interspaces between saddles are cream-yellow (in life); venter with heavy dark markings, each plate decorated by, anteriorly, a large rounded cone on either side joined to a smaller rounded cone in the centre; uniform phase pale grey-brown above, cream below; inter-prefrontal sulcus usually faintly marked; frontal usually marginally pitted (States 1 or 2); ventrals 226–247 in males, 241–271 in females ………………………………………………………………… D. abyssina
7b. Dorsal markings (saddles) usually not oblique, variable in shape (e.g. round, oval, square, rectangular), usually not confluent or forming zig-zags; no yellowish markings; venter varies from immaculate to heavily-marked, but not as extensively as above; inter-prefrontal sulcus variable; frontal pitting variable; ventrals 185–229 in males, 202–249 in females ……………………………………………………………………………………………………………… 8
8a. Dorsal markings (saddles) variable, often oval to rectangular (antero-posteriorly); pattern cycles 46–94 (mean 67.6); markings on venter variable; ventrals 185–226 in males, 202–249 in females ………………………………………………………………………………………… D. scabra
8b. Dorsal markings (saddles) large, often broad or elongate; pattern cycles 38–59; venter usually unmarked except for a few dark spots laterally ……………………………………………. 9
9a. Dorsal markings (saddles) cranio-caudally elongate, well separated, usually bifurcated at each end with a shallow but distinct notch; flanks with spots or blotches; ventrals 213–229 in males, 215–234 in females ………………………………………………………………… D. bazi
9b. Dorsal markings (saddles) broad, often squarish, usually separated by narrow sharply-defined white crossbars; flanks with elongate, wavy markings; ventrals 196–216 in males, 208–222 in females ………………………………………………………….. D. taylori sp. nov.
Acknowledgements
We thank the following museums and their curators and collection managers for providing access to specimens: Natural History Museum, London (C.J. McCarthy, P. Campbell); Museum National d’Histoire Naturelle, Paris (I. Ineich, N. Vidal); Museum für Naturkunde, Berlin (R. Günther, F. Tillack); Naturhistorisches Museum, Vienna (Silke Schweiger); Musée Royal de l’Afrique Centrale, Tervuren (D. Meirte); Museum of Comparative Zoology, Harvard (J.P. Rosado); Field Museum of Natural History, Chicago (A. Resetar); American Museum of Natural History, New York (C.W. Myers, D. Kizirian, D.A. Dickey); United States National Museum, Washington (G.R. Zug); University of Michigan Museum of Zoology, Ann Arbor (A.G. Kluge); California Academy of Sciences, San Francisco (R.C. Drewes, J.V. Vindum); Addis Ababa University Museum (G. Redda); Makerere University Zoological Museum, Kampala (R. Kityo); National Museums of Kenya, Nairobi (P.K. Malonza); Port Elizabeth Museum/Bayworld (W.R. Branch, W. Conradie); and KwaZulu-Natal Museum, Pietermaritzburg (C. Stoffels).
We thank the curators of the following museums for sending us material on loan: Los Angeles County Museum (R. Bezy); Museum Alexander Koenig, Bonn (W. Böhme); Muséum d’Histoire Naturelle, Geneva (A. Schmitz); National Museum of Kenya, Nairobi (P.K. Malonza); Naturhistorisches Museum, Vienna (H. Grillitsch); and Statens Naturhistoriske Museum, Københavns Universitet, Copenhagen (J.B. Rasmussen); and the curators and other individuals associated with the following museums for providing us with meristic data and/or images of specimens in their care: Museo Civico di Storia Naturale, Genova (G. Doria, M. Petri); Museo Civico di Storia Naturale, Milan (S. Scali); Museo di Storia Naturale, Università di Pavia, Pavia (E. Razzetti); Museo Zoologico “La Specola”, Università di Firenze (A. Nistri); Museo Regionale di Scienze Naturali di Torino, Turin (F. Andreone); Museum Alexander Koenig, Bonn (W. Böhme, M. Flecks, U. Bott); Statens Naturhistoriske Museum, Københavns Universitet, Copenhagen (J.B. Rasmussen); Institut Royal des Sciences Naturelles de Belgique, Brussels (G. Lenglet); National Museum of Kenya, Nairobi (P.K. Malonza); Port Elizabeth Museum/Bayworld (W.R. Branch, W. Conradie); Natural History Museum, London (P. Campbell); Museum National d’Histoire Naturelle, Paris (I. Ineich); Zoologisches Museum der Universität, Berlin (F. Tillack), Museum Alexander Koenig, Bonn (W. Böhme and U. Bott); California Academy of Sciences, San Francisco (J. Vindum); and Texas A&M University, Texas (T. Hibbits).
We also thank B. Hughes for providing scalation data for Ugandan specimens in the collection of the World Museum, Liverpool; the following individuals for photographs of Dasypeltis (see figures for credits): W.R. Branch, F. Finke, E. Greenbaum, T. Mazuch, S. Spawls, C. Tilbury and V. Trailin; V. Trailin for additional photographic records of D. scabra and D. atra in Ethiopia; S. Spawls for additional photographic records of D. atra and D. scabra in Kenya, and D. atra in Tanzania; T. Mazuch for additional photographic records of D. scabra and D. atra in Ethiopia; J. Marais for photographic records of Kenyan D. medici (Malindi) and D. atra (Emakoko); T. Dudu for photographic records of D. atra from Serengeti in Tanzania; C. Doria for a photographic record of a melanistic D. atra from Oldeani in Tanzania; A. Bauer, T. Mazuch, F. Tillack, D. Meirte, C. Kucharzewski, M.-O. Rodel, I. Ineich and R. Charles for access to old literature; S. Spawls for helping to trace various localities; and U. Ziliani for helping to trace records of D. scabra from Turkana County in Kenya. Frank Tillack provided collection details for Egyptian specimens in the collection in Berlin.
The first author’s trip to the natural history museums in London, Paris and Berlin in September and October 2011 was funded by the Reptile Speciation Project (a South African National Research Foundation [NRF] grant to K.A. Tolley, South African National Biodiversity Institute, Cape Town), his visit to the Royal Museum for Central Africa in Tervuren in October 2012 was made possible by an African Biodiversity Information Centre scholarship, and a later trip (September/October 2015) to the museum in Paris was funded by the National Museum (Bloemfontein).
We are indebted to J. Kersten (Bulawayo) and M. van Rensburg (National Museum, Bloemfontein) for preparing the distribution map; Zhongning Zhao (University of the Free State, Bloemfontein) for assistance with multivariate analyses; and W.R. Branch (Port Elizabeth Museum) for commenting on parts of the Discussion. The reviewers, Prof. A.M. Bauer (Villanova University, Philadelphia) and S. Spawls (Norwich, England) are thanked for their valuable comments and suggestions for improving the manuscript.
References
Anderson, J. 1896. A contribution to the herpetology of Arabia, with a preliminary list of the reptiles and batrachians of Egypt. London, 122 pp.
Anderson, J. 1898. Zoology of Egypt: Volume First. Reptilia and Batrachia. Bernard Quaritch, London, pp. i–lxv, 1–371.
Angel, F. 1925. Voyage de Ch. Allaud et R. Jeannel en Afrique Orientale (1911–12) Résultats scientifiques. Reptiles et batraciens. P. Lechevalier, Paris, 63 pp.
ANON. 2017. “Emin Pasha”. Wikipedia. (http://en.wikipedia.org/wiki/Emin_Pasha , accessed: 31 October 2017).
Baha El Din, S.M. 2006. A Guide to the Reptiles and Amphibians of Egypt. The American University in Cairo Press, Cairo, 359 pp.
Balletto, E. 1965. Contributo alla biogeographia della Somalia. Bollettino dei Musei e degli Istituti Biologici dell’Universita di Genova 36 (248): 191–280.
Bates, M.F. 2009. The egg-eating snakes of Africa. Culna 64: 2–4.
Bates, M.F. 2013. First records of the egg-eating snake Dasypeltis confusa Trape & Mané, 2006 in Nigeria and Chad, with range extensions for Ghana. Navorsinge van die Nasionale Museum, Bloemfontein 29 (2): 17–27.
BATES, M.F. 2017. Dasypeltis atra Sternfeld, 1912 (Reptilia: Serpentes): proposed precedence over Dasypeltis lineolata W. Peters, 1878. Bulletin of Zoological Nomenclature (submitted, Case 3750).
Bates, M.F. & Broadley, D.G. 2008a. Geographical Variation in Egg-eating Snakes of the Genus Dasypeltis in Africa south of Latitude 12° S. Sixth World Congress of Herpetology, Manaus, Brazil (17–22 August 2008). Available on CD with conference abstracts.
Bates, M.F. & Broadley, D.G. 2008b. Geographical variation in scalation and colour pattern in the egg-eating snakes of the genus Dasypeltis in Africa south of Latitude 12° S. Ninth Conference of the Herpetological Association of Africa, Sterkfontein Dam, Harrismith, South Africa (26–30 November 2008). Published in Abstract booklet.
Bates, M.F. & Ineich, I. 2012. A new size record for the West African egg-eating snake Dasypeltis gansi Trape & Mané, 2006, with new distribution records. Navorsinge van die Nasionale Museum, Bloemfontein 28 (3): 41–48.
Bates, M.F., Branch, W.R., Bauer, A.M., Burger, M., Marais, J., Alexander, G.J. & de Villiers, M.S. (Eds). 2014. Atlas and Red List of the Reptiles of South Africa, Lesotho and Swaziland. Suricata 1. South African National Biodiversity Institute, Pretoria, pp. i–xvii + 1–485.
BATES, M.F., BROADLEY, D.G., BARLOW, A., WüSTER, W. & TOLLEY, K.A. 2009. Egg-eating snakes of the genus Dasypeltis in Southern Africa: how many species are there? Abstracts: 50th Anniversary Conference of the Zoological Society of Southern Africa, Illovo Beach, South Africa, 21-25 July 2009. Pp. 69–70.
Bates, M.[F.], Broadley, D.[G.], Barlow, A., Wüster, W. & Tolley, K.[A.] 2012. Taxonomy and distribution of the African egg-eating snakes of the genus Dasypeltis. Abstracts: Seventh World Congress of Herpetology, Vancouver, Canada, 8–14 August 2012. African Herp News 58: 27–28.
BEHANGANA, M. & GOODMAN, J.D. 2002. Key to genera of snakes of Uganda (with a checklist and vernacular names). The Uganda Journal 48: 33–70.
Bianconi, J.J. 1850–1862. Specimina Zoologica Mosambicana. Bononiae Fascimile 1–16, 406 pp. [Fascimile 14 (1859), pp. 277–278, table 14]
Blanford, W.T. 1870. Observations on the geology and zoology of Abyssinia made during the progress of the British expedition to that country in 1867–1868. London, xii + 487 pp.
Bogert, C.M. 1940. Herpetological results of the Vernay Angola Expedition with notes on African reptiles in other collections. Part I – Snakes, including an arrangement of African Colubridae. Bulletin of the American Museum of Natural History (New York) 77 (1): 1–107.
Bosellini, A. 1986. East Africa continental margins. Geology 14: 76–78.
Boulenger, G.A. 1894. Catalogue of the snakes in the British Museum (Natural History), 2, containing the conclusion of the Colubridae Aglyphae. London, xi + 382 pp.
Boulenger, G.A. 1896a. A list of the reptiles and batrachians collected by Dr. Ragazzi in Shoa and Eritrea. Annali del Museo Civico di Storia Naturale Giacomo Doria (Genova), (Ser. 2) 16: 545–554.
Boulenger, G.A. 1896b. Report on Capt. Bottego’s second collection of reptiles and batrachians from Somaliland. Annali del Museo Civico di Storia Naturale Giacomo Doria (Genova), (Ser. 2) 17: 15–23.
Boulenger, G.A. 1896c. Catalogue of the snakes in the British Museum (Natural History), 3, containing the Colubridae (Opisthoglyphae and Proteroglyphae), Amblycephalidae and Viperidae. London, xi + 382 pp.
Boulenger, G.A. 1897. A list of the reptiles and batrachians of Somaliland and Gallaland. Annali del Museo Civico di Storia Naturale Giacomo Doria (Genova), (Ser. 2) 17: 275–280.
Boulenger, G.A. 1898. Concluding report on the late Capt. Bottego’s collection of reptiles and batrachians from Somaliland and British East Africa. Annali del Museo Civico di Storia Naturale Giacomo Doria (Genova), [1897–1898] (Ser. 2) 18: 715–723, Pls 9–10.
Boulenger, G.A. 1905. Descriptions of three new snakes discovered in South Arabia by Mr. G.W. Bury. Annals and Magazine of Natural History (London), (Ser. 7) 16 (93): 178–180.
Boulenger, G.A. 1909. Collezioni zoologische fatte nell’ Uganda dal Dott. E. Bayon. III. On a second collection of reptiles, batrachians and fishes made by Dr. E. Bayon in Uganda. Annali del Museo Civico di Storia Naturale Giacomo Doria (Genova), (Ser. 3) 4: 302–307.
Boulenger, G.A. 1911. Collezioni zoologische fatte nell’ Uganda dal Dott. E. Bayon. XIV. On a third collection of reptiles, batrachians and fishes made by Dr. E. Bayon in Uganda. Annali del Museo Civico di Storia Naturale Giacomo Doria (Genova), (Ser. 3) 5: 161–169.
Boulenger, G.A. 1912. Missione per la frontiera Italo-Etiopica sotto il Comando del Capitano Carlo Citerni. Risultati Zoologici. List of the reptiles and batrachians. Annali del Museo Civico di Storia Naturale Giacomo Doria (Genova), (Ser. 3) 5: 329–332.
Bourgeois, M. 1968. Contribution à la morphologie comparée du crâne des ophidiens del’Afrique centrale. Publications de l’Université Officielle du Congo à Lubumbashi 18: 1–293.
Branch, B. [W.R.] 2005. A Photoguide to Snakes other reptiles and amphibians of East Africa. Struik Publishers, Cape Town, 144 pp.
Branch, B. [W.R.] 2015. A Photoguide to Snakes other reptiles and amphibians of East Africa. Revised edition. Struik Nature (Random House Publishers), Cape Town, 160 pp.
Broadley, D.G. 1983. FitzSimons’ Snakes of Southern Africa. Delta Books, Johannesburg, 376 pp.
Broadley, D.G. 1990. FitzSimons’ Snakes of Southern Africa. Revised edition with addendum. Delta Books, Johannesburg, 376 pp. + Addendum.
Broadley, D.G. 1998. Introduction to the Reptilia, pp. ix–xxxiii. In: Contributions to the herpetology of the Belgian Congo based on the collection of the American Museum Congo Expedition, 1909–1915. Part I. Turtles, crocodiles, lizards and chameleons. Part II. Snakes. By Karl Patterson Schmidt. Reprint: Society for the Study of Reptiles and Amphibians, Ithaca.
Broadley, D.G. & Bates, M.F. 2009. New range extensions for Dasypeltis atra Sternfeld in Tanzania (Serpentes: Colubridae), with a review of the distribution of colour phases. African Journal of Herpetology 58 (1): 50–55.
Broadley, D.G., Doria, C.T. & Wigge, J. 2003. Snakes of Zambia. Chimaira, Frankfurt am Main, 280 pp.
Broadley, D.G. & Howell, K.M. 1991. A checklist of the reptiles of Tanzania, with synoptic keys. Syntarsus (Bulawayo) 1: 1–70.
Broadley, D.G. & Wallach, V. 2009. A review of the eastern and southern African blind-snakes (Serpentes: Typhlophidae), excluding Letheobia Cope, with the description of two new genera and a new species. Zootaxa 2255: 1–100.
Calabresi, E. 1925. Anfibi e rettili raccolti dal Signor Ugo Ignesti nell’Abissinia settentrionale. Atti della Sociatà Italiana di Scienze Naturali e dei Museo Civico di Storia Naturale in Milano 62 (2): 100–118.
Calabresi, E. 1927. Anfibi e rettili raccolti nella Somalia dal Proff. G. Stefanini e N. Puccioni (Gennaio-Luglio 1924). Atti della Sociatà Italiana di Scienze Naturali e dei Museo Civico di Storia Naturale in Milano 66 (1–2): 14–60.
Cherlin, V.A. 1990. Taxonomic revision of the snake genus Echis (Viperidae). II. An analysis of taxonomy and description of new forms. Proceedings of the Zoological Institute, Leningrad 207: 193–223.
CHISHOLM, H. (Ed.). 1911. “Emin Pasha”. Encyclopaedia Britannica 9. Eleventh Edition, Cambridge University Press, Cambridge (https://en.wikisource.org/wiki/1911_Encyclop%C3%A6dia_Britannica/Emin_Pasha , accessed: 31 October 2017).
Corkill, N.L. 1935. Notes on Sudan snakes: a guide to the species represented in the collection in the Natural History Museum, Khartoum. Sudan Government Natural History Museum, Khartoum, 40 pp.
Corkill, N.L. & Cochran, J.A. 1965. The snakes of the Arabian peninsula and Socotra. Journal of the Bombay Natural History Society 62 (3): 474–506.
Curry-Lindahl, K. 1956. Ecological studies on mammals, birds, reptiles and amphibians in the eastern Belgian Congo. Part I (Report no. 1 of the Lund University Congo Expedition 1951–1952). Annales du Musée Royal du Congo Belge (Tervuren), (8vo, Sciences Zoologiques) 42: 1–78.
Deane, J.B. 1833. The worship of the serpent traced throughout the world; attesting the Tempation and Fall of Man by the instrumentality of a Serpent Tempter. Second edition. Gilbert and Rivington, London, pp. i–xiv, 1–475.
DE QUIEROZ, K. 1998. The General Lineage Concept of Species, Species Criteria, and the Process of Speciation. A Conceptual Unification and Terminological Recommendations. Pp. 55-75. In: Howard, D.J. & Berlocher, S.H. (eds), Endless forms: species and speciation. Oxford University Press, U.S.A.
DE QUIEROZ, K. 2007. Species Concepts and Species Delimitation. Systematic Biology 56 (6): 879–886.
De Waal, S.W.P. 1978. The Squamata (Reptilia) of the Orange Free State, South Africa. Memoires van die Nasionale Museum, Bloemfontein 11: i–iii + 1–160.
De Witte, G.-F. 1962. Genera des Serpents du Congo et du Ruanda-Urundi. Annales de Musée Royal de l’Afrique Centrale (Tervuren), (8vo, Sciences Zoologiques) 104: 1–203 + 215 pls.
De Witte, G.-F. 1975. Boidae, Colubridae, Elapidae et Viperidae. In: Serpents. Exploration du Parc National des Virunga (2) 24: 62–121.
Dobiey, M. & Vogel, G. 2007. Venomous Snakes of Africa. Terralog 15, Chimaira, Frankfurt am Main., pp. 1–148 + foldout.
Duméril, A.M.C., Bibron, G. & Duméril, A.H.A. 1854. Erpétologie générale ou histoire naturelle compléte des Reptiles, Imprimerie Roret, Paris, vol. 7, part 1, xvi + 780 pp.
Emmrich, D. 1985. Bemerkungen und daten zur verbreitung eineger schlangenarten jn Athiopien. Aquarien Terrarien 32 (85): 65–68, 100–104, 136–140.
Emmrich, D. 1986. Die Afrikanische Eierschlange – Dasypeltis scabra. Aquarien Terrarien 33 (86): 208–211, 241–244, 277–281, 317–319, 350–354, 356.
Emmrich, D. 1992. Remarks on the distribution of some snake species of Ethiopia. Walia 14: 16–22.
Escoriza, D. 2010. Ecological niche modeling of two Afrotropical snakes: is the Sahara desert a true barrier for these species? Revista Española Herpetología 24: 93-100.
Escoriza, D. 2014. Dasypeltis sahelensis Trape & Mané, 2006 in Morocco and Western Sahara. Available from: http://www.moroccoherps.com/en/ficha/Dasypeltis_sahelensis/ (Date of access: 3 September 2014).
FIGUEROA, A., McKELVY, A.D, GRISMER, L.L., BELL, C.D. & LAILVAUX, S.P. 2016. A Species-Level Phylogeny of Extant Snakes with Description of a New Colubrid Subfamily and Genus. PLoS ONE 11 (9): e0161070.doi:10.1371/journal.pone.0161070.
Fischer, E. & Hinkel, H. 1992. Natur Ruandas/La Nature du Rwanda. Université Johannes Gutenberg (Institut de Botanique spéciale & Institut de Zoologie) et l’Institut de Recherche Scientifique et Technologique, Butare, 452 pp.
Fischer, J.G. 1884a. Über einige africanische Reptilien, Amphibien und Fische des Naturhistorischen Museums. I. Über die von Herrn Dr. G.A. Fischer im Massai-Gebiete (Ost-Afrika) auf seiner in Veranlassung der Geographischen Gesellschaft in Hamburg unternommenen Expedition gesammelten Reptilien, Amphibien und Fische. Jahrbuch Hamburgen Wissenschaftliche Anstalten 1: 1–32.
Fischer, J.G. 1884b. Herpetologische Bemerkungen. Abhandlungen aus dem Gebiete der Naturwissenschaften, Hamburg 8 (2): 43–51 (alternative page numbering: 3–11).
FitzSimons, V.F. 1962. Snakes of Southern Africa. Purnell & Sons, Cape Town, 423 pp.
Flemming, A.F. & Bates, M.F. 1999. Reptiles of Mkomazi. Pp. 411–428. In: Coe, M., McWilliam, N., Stone, G. & Packer, M. (Eds), Mkomazi: The ecology, biodiversity and conservation of a Tanzanian savanna. Royal Geographical Society, London, pp. i–viii + 1–608 + map.
FLOWER, S.S. 1933. Notes on the Recent Reptiles and Amphibians of Egypt, with a List of the Species recorded from that Kingdom. Proceedings of the Zoological Society of London. Pp. 735–851.
FROST, D.R. & HILLIS, D.M. 1990. Species in concept and practice: Herpetological applications. Herpetologica 46 (1): 87–104.
Gans, C. 1957. Dasypeltis medici lamuensis, a new race of egg-eating snake (Ophidia, Reptilia) from coastal East Africa. Breviora 79: 1–13.
Gans, C. 1959. A taxonomic revision of the African snake genus Dasypeltis (Reptilia : Serpentes). Annales du Musée Royal du Congo Belge (Tervuren), (8vo, Sciences zoologiques) 74: 1–237.
Gans, C. 1961. Mimicry in procryptically colored snakes of the genus Dasypeltis. Evolution 15 (1): 72–91.
Gans, C. 1964. Further comments on the forms of the African snake genus Dasypeltis (Reptilia : Serpentes). Revue de Zoologie et de Botanique Africaines 69 (3–4): 279–295.
Gans, C. 1971. Beobachtungen an afrikanischen Eierschlangen. Natur und Museum [1970] 100 (10): 460–471.
Gans, C. 1974. Biomechanics: An Approach to Vertebrate Biology. Lippincott, Philadelphia, i–x, 261 pp.
Gans, C. & Laurent, R.F. 1965. IV. Snakes. In: Gans, C., Laurent, R.F. & Pandit, H. Notes on a herpetological collection from the Somali Republic. Annales du Musée Royal de L’Afrique central (Tervuren), (8vo, Sciences zoologiques) 134: 47–70.
Gartner, G.E.A. & Greene, H.W. 2008. Adaptation in the African egg-eating snake: a comparative approach to a classic study in evolutionary functional morphology. Journal of Zoology, London 275: 368–374.
Gasco, F. 1876. Viaggio in Egitto dei socii P. Panceri e F. Gasco nell’anno 1873–1874. Parte seconda. Ritorno da Assuan. L’oasi del Fayum. Escursioni nei dintorni del Cairo. Caccia sul Lago Menzaleh. Annuario dell’ Associazione dei Naturalisti e Medici per la mutua Istruzione, Napoli, 1876: 1–146.
Gasperetti, J. 1988. Snakes of Arabia. Fauna of Saudi Arabia 9: 169–450.
GENIEZ, P. 2015. Serpents d’Europe, d’Afrique du Nord et du Moyen-Orient. Delachaux et Niestle, Paris, 380 pp.
Geniez, P. & Guillod, M. 2003. Status and new records of Dasypeltis scabra (Linnaeus, 1758), in Morocco. Herpetozoa 16 (1/2): 88–91.
GÖTHEL, H. 2015a. Die Pazifisten unter den Schlangen – Afrikanische Eierschlangen der Gattung Dasypeltis. Draco 16 (61): 6–21.
GÖTHEL, H. 2015b. Abgepacktes Futter – Vogeleier als Nahrungsquelle. Draco 16 (61): 22–27.
GÖTHEL, H. 2015c. Das Fressverhalten Afrikanische Eierschlangen. Draco 16 (61): 28–37.
GÖTHEL, H. 2015d. Mimikry und Abwehrverhalten bei Eierschlangen. Draco 16 (61): 46–52.
GÖTHEL, H. 2015e. Afrikanische Eierschlangen in Terrarium. Draco 16 (61): 54–61.
GÖTHEL, H. 2015f. Nachzucht von Afrikanischen Eierschlangen. Draco 16 (61): 62–69.
GÖTHEL, H. 2015g. Erkrankungen und Parasiten bei Eierschlangen. Draco 16 (61): 70–76.
GÖTHEL, H. 2015h. Die Eierschlangenarten Afrikas. Draco 16 (61): 78–92.
Gravlund, P. 2001. Radiation within the advanced snales (Caenophidia) with special emphasis on African opistoglyph colubrids, based on mitochondrial sequence data. Biological Journal of the Linnaean Society 72: 99–114.
Gravlund, P., Berggreen, A. & Andersen, M. 1998. Geographical distribution: Dasypeltis atra. Herpetological Review 29 (1): 53.
Günther, A. 1878. On reptiles from Midian collected by Major Burton. Proceedings of the Zoological Society of London, pp. 977–978.
Günther, A.L.G. 1894. Report on the collection of reptiles and fishes made by Dr. W. Gregory during his expedition to Mount Kenia. Proceedings of the Zoological Society of London, pp. 84–91.
Hinkel, H. 1986. Reptilien Ruandas. Mainz, 28 pp.
Hinkel, H. & Fischer, E. 1988. Reptiles et Amphibiens du Rwanda et leurs Environnement. Edition Bureau de Coordination, Kigali, Rwanda, 48 pp + 20 pls.
Hinkel, H. & Fischer, E. 1990. Checklist of amphibians and reptiles of Nyungwe Forest, Rwanda. Pp. 135–138. In: Dowsett, R.J. (Ed.), Enquête faunistique et floristique dans la Forêt de Nyungwe, Rwanda [Survey of the fauna and flora of Nyungwe Forest, Rwanda]. Tauraco Research Report 3.
Hoevers, L.G. & Johnson, P.M. 1982. Notes on a collection of snakes from the middle Juba region, Somalia. Monitore zoologico italiano, N.S. Supplement xvi (8): 173–203.
Hoser, R. 2013. A new egg-eating snake from the southern Arabian Peninsula (Squamata: Serpentes: Colubridae: Colubrinae: Boigini). Australasian Journal of Herpetology 21: 59–63.
Hughes, B. 1997. Dasypeltis scabra and Lamprophis fuliginosus – two pan-African snakes in the Horn of Africa: a tribute to Don Broadley. African Journal of Herpetology 46 (2): 68–77.
Ibrahim, A.A. 2013. The Herpetology of the Suez Canal Zone, Egypt. Vertebrate Zoology 63 (1): 87–110.
INEICH, I. I. 1999. Reptiles & Amphibiens de la République de Djibouti. Bilan des connaissances à l’aube du Troisième millénaire. Bureau National de la Diversitè Biologique (BNDB), Union International Pour la Conservation de la Nature (UICN), Programme des Nations-Unies pour le Développement (PNUD), Djibouti & Muséum National d’Histoire Naturelle (MNHN), Paris, pp. 1–60.
ICZN [International Commission on Zoological Nomenclature]. 1999. International Code of Zoological Nomenclature. Fourth Edition. International Trust for Zoological Nomenclature, London, XXIX + 306 pp.
IUCN [International Union for Conservation of Nature]. 2012. IUCN Red List Categories and Criteria: Version 3.1. Second edition. IUCN, Gland, Switzerland and Cambridge, UK.
IUCN [International Union for Conservation of Nature] [Standards and Petitions Subcommittee]. 2016. Guidelines for Using the IUCN Red List Categories and Criteria. Version 12. Prepared by the Standards and Petitions Subcommittee.
Downloadable from: http://www.iucnredlist.org/documents/RedListGuidelines.pdf.
Jan, G. 1863. Elenco Sistematico degli ofidi descritti e disegnati per l’Iconographia generale. Tipografia di A. Lombardi, Milano, viii + 143 pp.
Kaiser, H. 2014. Best Practices in Herpetological Taxonomy: Errata and Addenda. Herpetological Review 45 (2): 257–268.
Kaiser, H., Crother, B.I., Kelly, C.M.R., Luiselli, L., O’Shea, M., Ota, H., Passos, P., Schleip, W.D. & Wüster, W. 2013. Best Practices: In the 21st Century, Taxonomic Decisions in Herpetology are Acceptable Only When Supported by a Body of Evidence and Published via Peer-Review. Herpetological Review 44 (1): 8–23.
KELLY, C.M.R., BARKER, N.P. & VILLET, M.H. 2003. Phylogenetics of Advanced Snakes (Caenophidia) Based on Four Mitochondrial Genes. Systematic Biology 52 (4): 439–459.
Khalifa, M.A. & El-Khateeb, S.M. 2011. Fayoum Oasis between problems and potentials: Towards enhancing ecotourism in Egypt. Urbenviron Cairo 2011: 4th International Congress on Environmental Planning and Management. Green Cities: A Path to Sustainability. December 10 – 13, 2011. Cairo and El-Gouna, Egypt, pp. 1–16.
Kuper, R. & Kröpelin, S. 2006. Climate-Controlled Holocene Occupation in the Sahara: Motor of Africa’s Evolution. Science 313: 803–807.
Lanza, B. 1972. The reptiles of the Awash National Park (Shoa, Ethiopia). Monitore Zoologico Italiano. Supplement 4 (7): 159–184.
Lanza, B. 1983. A list of the Somali amphibians and reptiles. Monitore Zoologico Italiano (Firenze), (Ser. 2), Supplement 18 (8): 193–247.
Lanza, B. 1990. Amphibians and reptiles of the Somali Democratic Republic: checklist and biogeography. Biogeographia (Bologna), (1988) 14: 407–465.
Largen, M.J. 1997. An annotated checklist of the amphibians and reptiles of Eritrea, with keys for their identification. Tropical Zoology (Firenze) 10: 63–115.
Largen, M.J. & Rasmussen, J.B. 1993. Catalogue of the snakes of Ethiopia (Reptilia Serpentes), including identification keys. Tropical Zoology (Firenze) 6 (2): 313–434.
Largen, M.J. & Parker, A.R. 2004. Catalogue of the spirit-preserved herpetological collections in the Liverpool Museum. National Museums, Liverpool, 47 pp.
Largen, M. & Spawls, S. 2010. The Amphibians and Reptiles of Ethiopia and Eritrea. Chimaira, Frankfurt am Main, 693 pp.
Laurent, R.F. 1956. Contribution à l’Herpetologie de la région des Grands Lacs de l’Afrique centrale. I: Généralitiés—II. Chéloniens – III. Ophidiens. Annales du Musée Royal du Congo Belge (Tervuren), (8vo, Sciences Zoologiques) 48: 1–390, pls. 1–31.
Laurent, R.F. 1960. Notes complémentaires sur les Chélonians et les Ophidiens du Congo oriental. Annales du Musée Royal du Congo Belge (Tervuren), (8vo, Sciences Zoologiques) 84: 1–86.
Laurent, R.F. 1965. Contribution à l’histoire de l’herpetologie congolaise et bibliographie generale. Academie Royal des Sciences d’Outre Mer. Classe des Sciences naturelles et médicales (N.S.) 16: 1–55.
Lavin, B.R. & Papenfuss, T.J. 2012. The phylogenetic position of Chalcides ocellatus (Squamata: Scincidae) from Yemen and Somalia. Zootaxa 3221: 26–36.
LAWSON, R., SLOWINSKI, J.B., CROWTHER, B.I. & BURBRINK, F.T. 2005. Phylogeny of the Colubroidea (Serpentes): New evidence from mitochondrial and nuclear genes. Molecular Phylogenetics and Evolution 37: 581–601.
Leviton, A.E. 1986. Description of a new species of Coluber (Reptilia: Serpentes: Colubridae) from the southern Tihama of Saudi Arabia, with comments on the biogeography of southwestern Arabia. Fauna of Saudi Arabia 8: 436–446.
Leviton, A.E. & Aldrich, M.L. 1984. John Anderson: a zoologist in the Victorian period, pp. v–xxxv. In: K. Adler (Ed.), Herpetology of Arabia, by John Anderson. Reprint edition, Society for the Study of Amphibians and Reptiles, Athens, Ohio.
Lichtenstein, M.H.C. & VON MARTENS, C.E. 1856. Nomenclator reptilium et amphibiorum Musei Zoologici Berolinensis. Namenverzeichniss der in der zoologischen Sammlung der Königlichen Universität zu Berlin aufgestellten Arten von Reptilien und Amphibien nach ihren Ordungen, Familien und Gattungen. Buchdrukerei der Königlichen Akademie der Wissenschaften, Berlin, 48 pp.
Linnaeus, C. 1758. Systema naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, diferentiis, synonymis, locis. Holmia [Stockholm], Laurentii Salvii. Tenth edition. Vol. 1, 825 pp.
Lönnberg, E. 1922. Sammlungen der Schwedischen Elgon-Expedition im Jahre 1920. 6. Reptiles. Arkiv för Zoologi (Uppsala) 14 (12): 1–8.
Lönnberg, E. & Andersson, L.G. 1913. On a collection of reptiles from Kismayu. Arkiv för Zoologi (Uppsala) 8 (20): 1–6.
Loveridge, A. 1916. Report on the collection of Ophidia in the Society’s Museum. Journal of the East Africa and Uganda Natural History Society (Nairobi) 5 (10): 76–87.
Loveridge, A. 1929. East African reptiles and amphibians in the United States National Museum. Bulletin of the United States National Museum (Washington) (151): 1–135.
Loveridge, A. 1936a. African reptiles and amphibians in Field Museum of Natural History. Field Museum of Natural History Zoological Series (Chicago) 22 (1): 5–111.
Loveridge, A. 1936b. Scientific results of an expedition to rain forest regions in eastern Africa. V. Reptiles. Bulletin of the Museum of Comparative Zoology (Harvard) 79 (5): 209–337, Pls 1–9.
Loveridge, A. 1942. Scientific results of a fourth expedition to forested areas in East and Central Africa. Bulletin of the Museum of Comparative Zoology (Harvard) 91 (4): 237–373, Pls 1–6.
Loveridge, A. 1946. A guide to the snakes of the Nairobi district. Journal of the East Africa Natural History Society (Nairobi) 18 (3&4): 97–115.
Loveridge, A. 1956. On snakes collected in the Anglo-Egyptian Sudan by J.S. Owen, Esq. Sudan Notes and Records (Khartoum) (1955) 36: 37–56.
Loveridge, A. 1957. Check list of the reptiles and amphibians of East Africa (Uganda; Kenya; Tanganyika; Zanzibar). Bulletin of the Museum of Comparative Zoology (Harvard) 117 (2): 151–362 + i–xxxvi.
Malonza, P.K., Bwong, B.A. & Muchai, V. 2011. Kitobo Forest of Kenya, a unique hotspot of herpetofaunal diversity. Acta Herpetologica 6 (2): 149–160.
MÄNNEL, E. & KUCHARZEWSKI, C. 2017. Von exotischen Kaltblütern zum deutschen vollblut – Leben und Werk des Herpetologen und Hippologen RICHARD STERNFELD (1884 – 1943). Sekretär 17 (1): 3–50.
Marx, H. 1968. Checklist of the Reptiles and Amphibians of Egypt. Special Publication, United States Naval Medical Research Unit Number Three, Cairo, pp. i-iii, 1–91.
Mazuch, T. 2013. Amphibians and Reptiles of Somaliland and Eastern Ethiopia, based on two field trips in 2010 / 2011. Tomáš Mazuch, Dříteč, Czech Republic.
Meirte, D. 1992. Clés de determination des serpents d’Afrique. Annalen Zoologische Wetenschappen 267: 1–152.
Mertens, R. 1938. Über eine herpetologische Sammlung aus dem Gebiete des Njarasa-Grabens Ost-Afrika. Veröffentlichungen aus dem Deutschen Kolonial-und Übersee-Museum in Bremen 2 (1): 1–16.
Mertens, R. 1954. Neue Schlangenrassen aus Südwest- und Südafrika. Zoologischer Anzeiger, Leipzig 152 (9/10): 213–219.
Mertens, R. 1955. Die Amphibien und Reptilien Südwestafrikas. Aus den Ergebnissen einer im Jahre 1952 ausgeführten Reise. Abhandlungen der Senckenbergischen Naturforschenden Gesellschaft 490: 1–172.
MINTER, L.R., BURGER, M., HARRISON, J.A., BRAACK, H.H., BISHOP, P.J. & KLOEPFER, D. 2004. Atlas and Red Data Book of the Frogs of South Africa, Lesotho and Swaziland. SI/MAB Series no. 9. Smithsonian Institution, Washington D.C.
Mockrin, M. & Thieme, M. (Undated). Nile Delta flooded savanna. Available from: http://www.worldwildlife.org/ecoregions/pa0904 (Date of access: 11 June 2014).
Mocquard, F. 1888. Sur une collection de reptiles et batraciens rapports des Pays Çomalis et de Zanzibar par M.C. Révoil. Mémoires Publies par la Société Philomathique a l’Occasion du Centenaire de sa Fondation, pp. 109–134.
NeҪas, P. & Modry, D. 1998a. Commentarios sobre la biologia, matenimento y reproduccion de Dasypeltis atra. Reptilia 4: 22–27.
NeҪas, P. & Modry, D. 1998b. Remarks on the biology, keeping and breeding of the African egg-eating snake Dasypeltis atra. Reptilia, The European Herp Magazine 4: 22-27.
Panceri, P. 1878. Catalogo Sistematico del Gabinetto di Anatomia Comparata nella Regia Universita degli Studi di Napoli. Supplemento II. Tipografia dell’ Accademia Reale delle Scienze, Napoli, 55 pp.
Parker, H.W. 1932. Scientific results of the Cambridge expedition to the East African lakes 1930–1. 5. Reptiles and amphibians. Journal of the Linnean Society, London, Zoology 38 (258): 213–229.
Parker, H.W. 1949. The snakes of Somaliland and the Sokotra Islands. Zoologische Verhandelingen (Leiden), (6), 1–115.
Peracca, M.G. 1909. Rettili ed amfibii. Pp. 165–180. In: De Filippi, F. & Amedeo di Savoia, L. (Eds), Il Ruwenzori. Relazioni scientifiche. Resultati scientific della Spedizione Condotta da S.A.R. il Duca degli Abruzzi al Ruwenzori. Vol. I. Zoologia, botanica. Ulrico Hoepli Libraio della Real Casa, Milano.
Peters, W.C.H. 1864. Eine neue Art der Baumvipern, Atheris polylepis, aus Liberia. Monatsberichte der königliche Preussischen Akademie der Wissenschaften zu Berlin, pp. 642–645 (footnotes pp. 644–645).
Peters, W.C.H. 1878. Über die von Hrn. J.M. Hildebrandt während seiner letzen ostafrikanischen Reise gesammelten Säugethiere und Amphibien. Monatsberichte der königliche Preussischen Akademie der Wissenschaften zu Berlin 1878: 194–209.
Petzold, A., Vargas-Ramírez, M., Kehlmaier, C., Vamberger, M., Branch, W.R., Du Preez, L., Hofmeyr, M.D., Meyer, L., Schleicher, A., Široký, P. & Fritz, U. 2014. A revision of African helmeted terrapins (Testudines: Pelomedusidae: Pelomedusa), with descriptions of six new species. Zootaxa 3795 (5): 523–548.
Pitman, C.R.S. 1974. A Guide to the Snakes of Uganda. Revised edition. Wheldon & Wesley, Codicote, 289 pp.
RASMUSSEN, J.B. 2005. On the identification and distribution of the Two-Striped Night Adder (Causus bilineatus) and related forms. African Journal of Herpetology 54 (1): 1–15.
Rehak, I. & Osborn, D.J. 1988. Notes on the distribution of reptiles and amphibians in Egypt. Véstník Ceskoslovenské Společnosti Zoologické 52: 271–277.
Roux, J. 1910. Reptilien und Amphibien. Revue Suisse de Zoologie (Geneve) 18: 95–103.
SABAJ, M.H. 2016. Standard symbolic codes for institutional resource collections in herpetologyand ichthyology: an Online Reference. Version 6.5 (16 August 2016). American Society of Ichthyologists and Herpetologists, Washington, D.C. (http://www.asih.org/ , accessed 25 October 2017).
Saleh, M.A. 1997. Amphibians and Reptiles of Egypt. Publication of the Biodiversity Unit 6: 1–234.
SALEH, M. & SARHAN, M. 2016. The egg-eating snake (Colubridae: Dasypeltis) of Faiyum, Egypt, with the description of a new species. Bulletin de la Société Herpétologique de France 160: 25–48.
Schätti, B. & Desvoignes, A. 1999. The herpetofauna of southern Yemen and the Sokotra Archipelago. Museum d’Histoire naturelle Geneve, 178 pp., 40 pls.
Schätti, B. & Gasperetti, J. 1994. A contribution to the herpetofauna of southwest Arabia. Fauna of Saudi Arabia 14: 348–423.
Schätti, B. & Ineich, I. 2004. A new racer of the genus Platyceps Blyth from Djibouti (Reptilia: Squamata: Colubrinae). Revue suisse de Zoologie 111 (4): 685–690.
Schleich, H.H., Kästle,W. & Kabisch, K. 1996. Amphibians and Reptiles of North Africa. Koeltz, Koenigstein, 627 pp.
Schleip, W.D. 2014. Two New Species of Leiopython Hubecht, 1879 (Pythonidae: Serpentes): Non–compliance with the International Code of Zoological Nomenclature Leads to Unavailable Names in Zoological Nomenclature. Journal of Herpetology 48 (2): 272–275.
Scortecci, G. 1930. Rettili ed anfibi raccolti dal prof. E. Zavattari in Eritrea. Atti della Società Italiana si Scienze Naturali e del Museo Civico di Storia Naturale in Milano 9 (3–4): 193–217.
Scortecci, G. 1931. Terzo contributo alla conoscenza dei rettili della Somalie Italiana (Ofidi). Atti della Società Italiana si Scienze Naturali e del Museo Civico di Storia Naturale in Milano 70 (1): 203–215.
Scortecci, G. 1932. Rettili del Yemen. Atti della societa italiana di Scienze Naturali e del Museo Civico di Storia Naturale in Milano 69 (3–4): 319–321.
Scortecci, G. 1939. Spedizione zoological del Marchese Saverio Patrizi nel Basso Guiba e nell’Oltregiuba. Giugno-Agosto 1934 XII. Rettili. Ofidi. Annali del Museo Civico di Storia Naturale Giacomo Doria (Genova) 58: 263–291.
Scortecci, G. 1943. Reptilia. In: Missione Biologica Sagan-Omo, 7. Zoologia 1: 267-305.
SINDACO, R., VENCHI, A. & GRIECO, C. 2013. Reptiles of the Western Palearctic. 2. Annotated checklist and distributional atlas of the snakes of Europe, North Africa, Middle East and Central Asia, with update to the Vol. 1. Edizioni Belvedere, Latina (Italy), 543 pp.
Smith, A. 1838–1849. Illustrations of the zoology of South Africa, consisting chiefly of figures and descriptions of the objects of natural history collected during an expedition into the interior of South Africa, in the years 1834, 1835 and 1836, fitted out by ‘The Cape of Good Hope Association for Exploring Central Africa.’ Vol. III. Reptilia. Smith, Elder & Co., London, 191 pp., Pls. 1–78 + app.
Spano, S. 1972. Missioni Scortecci nell’Arabia Meridionale (1962–1965): Leptotyphlopidae e Colubridae (Ophidia). Bolletino di Museo di Istituto di Zoologia dell’Università di Genova 40: 131–135.
Spawls, S. 1978. A checklist of the snakes of Kenya. Journal of the East Africa Natural History Society and National Museum (Nairobi) 31 (167): 1–18.
Spawls, S. 2004. A New Species of Pseudoboodon (Reptilia: Serpentes) from the Central Highlands of Ethiopia; with notes on some other members of the genus. African Journal of Herpetology 53 (1): 13–19.
SPAWLS, S. 2017. A Guide to Amphibians and Reptiles of the Maasai Mara. Privately published: Stephen Spawls, 40 pp.
Spawls, S., Howell, K.M. & Drewes, R.C. 2006. Pocket guide to the reptiles and amphibians of East Africa. A.C. Black, London, 240 pp.
Spawls, S., Howell, K., Drewes, R. & Ashe, J. 2002. A field guide to the reptiles of East Africa (Kenya, Tanzania, Uganda, Rwanda and Burundi). Academic Press, San Diego, 543 pp.
Stejneger, L. 1894. On some collections of reptiles and batrachians from East Africa and the adjacent islands, recently received from Dr. W.L. Abbott and Mr. William Astor Chanler, with descriptions of new species. Proceedings of the United States National Museum (Washington) [1893] 16 (970): 711–741.
Sternfeld, R. 1908a. Zur Schlangenfauna Ostafrikas. I. Schlange aus Süd-Abessinien. Mittheilungen aus dem Zoologischen Museum in Berlin 4 (1): 239–241.
Sternfeld, R. 1908b. Zur Schlangenfauna Ostafrikas. II. Schlange aus Britisch-Ostafrika. Mittheilungen aus dem Zoologischen Museum in Berlin 4 (1): 242–245.
Sternfeld, R. 1912. Reptilia. Pp. 197–280, Pls VI–IX. In: Schubotz, H. (Ed.), Wissenschaftliche Ergebnisse der deutschen Zentral-Afrika-Expedition, 1907–1908. 4, Zoologie II, Klinkhardt & Biermann, Leipzig. [Entire volume published in 1913].
Trape, J.-F. & Mané, Y. 2006. Le genre Dasypeltis Wagler (Serpentes: Colubridae) en Afrique de l’Ouest: description de trois espèces et d’une sous-espèce nouvelles. Bulletin de Société Herpétologique de France 119: 27–56.
Trape, J.-F., Chirio, L., Broadley, D.G. & Wüster, W. 2009. Phylogeography and systematic revision of the Egyptian cobra (Serpentes: Elapidae: Naja haje) species complex, with the description of a new species from West Africa. Zootaxa 2236: 1–25.
Trape, S., Mediannikov, O. & Trape, J.-F. 2012. When colour patterns reflect phylogeography: new species of Dasypeltis (Serpentes: Colubridae: Boigini) from West Africa. Comptes Rendus Biologies 335: 488–501.
Uthmöller, W. 1937. Beitrag zur Kenntnis der Schlangenfauna des Kilimandscharo (Tanganyika-Territory, ehemaliges Deutsch-Ostafrika). Temminckia (Leiden) 2: 97–134.
Van As, J., du Preez, J., Brown, L. & Smit, N. 2012. The story of life and the environment: an African perspective. Struik Nature, Cape Town, 456 pp.
VAN DE LOOCK, D. & BATES, M.F. 2016. Dasypeltis atra, Montane Egg-eater: Diet and distribution: African Herp News 63: 16–18.
VATS, R. & SAFARI, I. 2014. Diversity of Snakes at the University of Dodoma Campus, Tanzania. American Journal of Zoological Research 2 (3): 41–45.
Venchi, A. & Sindaco, R. 2006. Annotated checklist of the reptiles of the Mediterranean countries, with keys to species identification. Part 2 – Snakes (Reptilia, Serpentes). Annali del Museo di Storia Naturale “G. Doria”, Genova 98: 259–364.
Vetaas, O.R., Salih, E.A. & Jurasinski, G. 2012. Vegetation changes in the Red Sea Hills: from mist oasis to arid shrub. Plant Ecology & Diversity 5 (4): 527–539.
VONESH, J. 2001. Natural history and biogeography of the amphibians and reptiles of Kibale National Park, Uganda. Contemporary Herpetology 2001 (4): 1–14.
WAGLER, J. 1830. Natürliches System der Amphibien, mit vorangehender Classification der Säugthiere und Vögel. Ein Beitrag zur vergleichenden Zoologie. Munchen. Pp. iii–vi, 1–354.
WAGNER, P. & BÖHME, W. 2007. Herpetofauna Kakamegensis – The amphibians and reptiles of Kakamega Forest, western Kenya. Bonner zoologische Beitrage 55 (2): 123–150.
WALLACH, V., WILLIAMS, K.L. & BOUNDY, J. 2014. Snakes of the World: A Catalogue of Living and Extinct Species. CRC Press, Boca Raton.
WALLACH, V., WÜSTER, W. & BROADLEY, D.G. 2009. 2009. In praise of subgenera: taxonomic status of cobras of the genus Naja Laurenti (Serpentes: Elapidae). Zootaxa 2236: 26-36.
Watkins-Colwell, G.J., Dornburg, A., Hawlena, D. & Moore, J.A. 2010. The Amphibians and Reptiles of the 1962–1965 Yale University Prehistoric Expedition to Nubia. Bulletin of the Peabody Museum of Natural History 51 (2): 179–197.
Wüster, W. & Broadley, D.G. 2003. A new species of spitting cobra (Naja) from north-eastern Africa (Serpentes: Elapidae). Journal of Zoology, London 259: 345–359.
Zavattari, E. 1930. Gli animali velenosi della Colonia Eritrea. In: Bono, S.E.E. – Ministro delle Colonie – della Missione Scientifica per l’Eritrea (Parma) 8: 171–205.
Ziliani, U., Sindaco, R., Razetti, E., Wasonga, V., Modrý, D., Necas, P., Carugati, C & Fasola, M. 2006. The herpetofauna of the Eastern side of the Lake Turkana (Northern Kenya). Pp. 192–193. In: Bologna, M.A., Capula, M., Carpaneto, G.M., Luiselli, L., Marangoni, C. & Venchi, A. Abstracts: 6° Congresso Nazionale della Societas Herpetologica Italica (Roma, 27 September – 1 October 2006), Stilografica, Roma.
Table 1. Numbers of pattern cycles and other pattern morphs in Dasypeltis from north-eastern Africa and south-western Arabia. Codes in parentheses under ‘Range’ and ‘Other morphs’ are those used by Gans (1959) for the various dorsal colour patterns.
| N | Range | Mean | S.D. | Other morphs |
| D. medici | 24 | 57–108 (8) | 74.0 | 11.9 | Faintly patterned to uniform (3): 37
Total patterned (8): 35 |
| D. confusa | 30 | 50–79 (5L) | 61.7 | 7.1 | Uniform brown: 11 |
| D. scabra | 107 | 46–94 (5N) | 67.4 | 10.0 | |
| D. bazi (Egypt) | 5 | 39–49 (5N) | 45.2 | 4.4 | |
| D. cf. bazi (Sudan) | 1 | 48 | |||
| D. taylori sp. nov. | 8 | 44–59 (5N) | 52.8 | 5.1 | |
| D. atra | 51 | 62–130 (2Bx) | 91.8 | 14.1 | Melanistic (2M): 89, Brown (2B): 91, Total 2Bx: 135 |
| D. fasciata | 5 | 108–141 (4) | 117.8 | 13.3 | |
| D. abyssina | 11 | 65–98 (5N, 5I) | 80.7 | 11.4 | Uniform brown/tan: 7 |
| D. crucifera sp. nov. | 3 | 51-62 (5I) | 55.0 | ||
| D. arabica sp. nov. | 6 | 62–79 (5I) | 70.0 | 5.7 |
Table 2. Ventral and subcaudal scale counts in Dasypeltis from north-eastern Africa and south-western Arabia. Values for males are in bold.
| VENTRALS SUBCAUDALS
Males Females Males Females |
||||||||||||||||
| N | Range | Mean | S.D. | N | Range | Mean | S.D. | N | Range | Mean | S.D. | N | Range | Mean | S.D. | |
| D. medici | 29 | 220–252 | 235.8 | 8.1 | 49 | 218–252 | 234.4 | 7.3 | 27 | 75–109 | 88.3 | 8.1 | 47 | 61–90 | 77.5 | 5.7 |
| D. confusa | 14 | 199–224 | 209.9 | 7.1 | 30 | 212–239 | 226.1 | 7.9 | 12 | 51–67 | 60.2 | 5.3 | 24 | 48–61 | 54.1 | 3.0 |
| D. scabra | 50 | 185–226 | 209.3 | 8.9 | 76 | 202–249 | 226.9 | 8.5 | 45 | 47–73 | 62.7 | 5.6 | 72 | 43–65 | 53.6 | 4.8 |
| D. bazi (Egypt) | 3 | 219–229 | 225.7 | 5.8 | 3 | 227–234 | 229.7 | 3.8 | 3 | 59–60 | 59.3 | 0.6 | 3 | 50–55 | 52.0 | 2.6 |
| D. cf. bazi (Sudan) | 1 | 229 | 1 | 67 | ||||||||||||
| D. taylori sp. nov. | 5 | 196–216 | 202.2 | 8.0 | 3 | 208–222 | 213.7 | 7.4 | 4 | 55–68 | 60.5 | 5.4 | 2 | 44–52 | 48.0 | 5.7 |
| D. atra | 109 | 199–232 | 212.7 | 6.8 | 167 | 211–256 | 225.7 | 7.9 | 103 | 50–71 | 61.9 | 3.9 | 151 | 45–64 | 53.5 | 3.3 |
| D. fasciata | 3 | 212–230 | 223.7 | 10.1 | 9 | 237–248 | 242.7 | 3.8 | 3 | 59–76 | 68.7 | 8.7 | 9 | 64–76 | 68.3 | 3.7 |
| D. abyssina | 6 | 226–247 | 235.5 | 8.2 | 10 | 241–271 | 256.0 | 9.3 | 4 | 57–68 | 61.5 | 4.6 | 9 | 49–65 | 56.7 | 5.0 |
| D. crucifera sp. nov. | 1 | 226 | 3 | 231–247 | 240.3 | 8.3 | 1 | 61 | 3 | 49–52 | 50.3 | 1.5 | ||||
| D. arabica sp. nov. | 3 | 236–244 | 239.3 | 4.2 | 4 | 239–254 | 244.8 | 6.9 | 3 | 63–65 | 63.7 | 1.2 | 4 | 53–61 | 56.8 | 3.5 |
Table 3. Loading scores for Principal Component Analysis of Dasypeltis males in north-eastern Africa and south-western Arabia.
| PC 1 | PC 2 | |
| Midbody scale rows | 0.2344 | –0.3343 |
| Ventrals | 0.3292 | 0.3966 |
| Subcaudals | 0.3438 | 0.5406 |
| Supralabials | 0.0471 | 0.0288 |
| Preoculars | 0.0193 | 0.2555 |
| Postoculars | 0.4007 | –0.2850 |
| Anterior temporals | 0.3447 | 0.0854 |
| Posterior temporals | 0.3890 | –0.2412 |
| Pattern cycles | 0.3316 | –0.2163 |
| Frontal pitting | 0.3194 | 0.2843 |
| Inter-prefrontal sulcus | 0.2706 | –0.3135 |
Table 4. Loading scores for Principal Component Analysis of Dasypeltis females in north-eastern Africa and south-western Arabia.
| PC 1 | PC 2 | |
| Midbody scale rows | 0.2960 | 0.2813 |
| Ventrals | 0.2792 | –0.0893 |
| Subcaudals | 0.3261 | –0.4928 |
| Supralabials | 0.1168 | 0.0920 |
| Preoculars | 0.1024 | 0.3277 |
| Postoculars | 0.4236 | 0.1670 |
| Anterior temporals | 0.3542 | –0.2803 |
| Posterior temporals | 0.4146 | –0.0816 |
| Pattern cycles | 0.2204 | 0.4892 |
| Frontal pitting | 0.3748 | –0.1808 |
| Inter-prefrontal sulcus | 0.1955 | 0.4113 |
Table 5. Observed (rows) and predicted (columns) classifications of male Dasypeltis from north-eastern Africa and south-western Arabia according to the Discriminant Function Analysis.
| % correct | D.
atra p = .430 |
D. scabra
p = .258 |
D. confusa
p = .065 |
D. medici
p = .097 |
D. fasciata
p = .022 |
D. taylori
p = .032 |
D. abyssina
p = .043 |
D. arabica
p = .022 |
D.
bazi p = .032 |
|
| D. atra | 85.0 | 34 | 5 | 0 | 0 | 0 | 0 | 1 | 0 | 0 |
| D. scabra | 83.3 | 2 | 20 | 1 | 0 | 0 | 0 | 0 | 0 | 1 |
| D. confusa | 50.0 | 1 | 2 | 3 | 0 | 0 | 0 | 0 | 0 | 0 |
| D. medici | 100 | 0 | 0 | 0 | 9 | 0 | 0 | 0 | 0 | 0 |
| D. fasciata | 100 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 |
| D. taylori | 66.7 | 0 | 1 | 0 | 0 | 0 | 2 | 0 | 0 | 0 |
| D. abyssina | 100 | 0 | 0 | 0 | 0 | 0 | 0 | 4 | 0 | 0 |
| D. arabica | 100 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 |
| D. bazi | 66.7 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 |
| Total | 83.9 | 38 | 28 | 4 | 9 | 2 | 2 | 5 | 2 | 3 |
Table 6. Standardised Canonical Discriminant Function Coefficients (Sigma-restricted parameterisation) for the analysis of male Dasypeltis in north-eastern Africa and south-western Arabia.
| Function 1 | Function 2 | |
| Midbody scale rows | –0.001839 | 0.229281 |
| Ventrals | 0.604129 | –0.405545 |
| Subcaudals | 0.812478 | 0.007712 |
| Supralabials | 0.137091 | –0.034382 |
| Preoculars | –0.006490 | –0.090852 |
| Postoculars | 0.099070 | 0.304857 |
| Anterior temporal | 0.007210 | –0.104399 |
| Posterior temporal | –0.075543 | 0.170241 |
| Pattern cycles | –0.085729 | 0.541770 |
| Frontal pitting | 0.190038 | 0.669032 |
| Inter-prefrontal sulcus | 0.020179 | 0.300686 |
| Eigenvalue | 6.032452 | 2.042220 |
| Cumulative Proportion | 0.591676 | 0.791982 |
Table 7. Observed (rows) and predicted (columns) classifications of female Dasypeltis from north-eastern Africa and south-western Arabia according to the Discriminant Function Analysis.
| % correct | D.
atra p = .400 |
D. scabra
p = .258 |
D. confusa
p = .084 |
D. medici
p = .129 |
D. fasciata
p = .019 |
D. taylori
p = .013 |
D. abyssina
p = .045 |
D. arabica
p = .026 |
D.
bazi p = .013 |
D. crucifera
p = .013 |
|
| D. atra | 74.2 | 46 | 6 | 6 | 0 | 0 | 0 | 3 | 1 | 0 | 0 |
| D. scabra | 75.0 | 2 | 30 | 3 | 0 | 0 | 2 | 0 | 2 | 1 | 0 |
| D. confusa | 61.5 | 3 | 2 | 8 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| D. medici | 95.0 | 1 | 0 | 0 | 19 | 0 | 0 | 0 | 0 | 0 | 0 |
| D. fasciata | 100 | 0 | 0 | 0 | 0 | 3 | 0 | 0 | 0 | 0 | 0 |
| D. taylori | 50.0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 |
| D. abyssina | 100 | 0 | 0 | 0 | 0 | 0 | 0 | 7 | 0 | 0 | 0 |
| D. arabica | 50.0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 |
| D. bazi | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| D. crucifera | 50.0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 |
| Total | 75.5 | 55 | 41 | 17 | 19 | 3 | 3 | 10 | 5 | 1 | 1 |
Table 8. Standardised Canonical Discriminant Function Coefficients (Sigma-restricted parameterisation) for the analysis of female Dasypeltis in north-eastern Africa and south-western Arabia.
| Function 1 | Function 2 | |
| Midbody scale rows | 0.155036 | –0.307790 |
| Ventrals | 0.174019 | 0.644477 |
| Subcaudals | 0.919766 | 0.182047 |
| Supralabials | 0.000409 | 0.009540 |
| Preoculars | –0.005440 | 0.252592 |
| Postoculars | 0.203638 | –0.274530 |
| Anterior temporal | 0.420286 | 0.032552 |
| Posterior temporal | –0.077620 | 0.051138 |
| Pattern cycles | –0.041610 | –0.269700 |
| Frontal pitting | 0.256583 | –0.673860 |
| Inter-prefrontal sulcus | –0.110640 | –0.282740 |
| Eigenvalue | 5.033508 | 1.395425 |
| Cumulative Proportion | 0.627753 | 0.801783 |
Table 9. Loading scores for Principal Component Analysis of Dasypeltis males in the Horn of Africa and Egypt.
| PC 1 | PC 2 | |
| Midbody scale rows | 0.3324 | –0.0988 |
| Ventrals | 0.1386 | –0.4114 |
| Subcaudals | 0.0951 | 0.1427 |
| Supralabials | 0.1531 | –0.3130 |
| Postoculars | 0.5229 | –0.1184 |
| Anterior temporal | 0.2389 | 0.0896 |
| Posterior temporal | 0.3864 | 0.1763 |
| Pattern cycles | 0.5052 | –0.1264 |
| Frontal pitting | 0.1129 | 0.6546 |
| Inter-prefrontal sulcus | 0.3001 | 0.4529 |
Table 10. Loading scores for Principal Component Analysis of Dasypeltis females in the Horn of Africa and Egypt.
| PC 1 | PC 2 | |
| Midbody scale rows | 0.4654 | –0.2276 |
| Ventrals | –0.3110 | 0.4003 |
| Subcaudals | 0.0721 | 0.2901 |
| Supralabials | –0.2623 | 0.3800 |
| Preoculars | 0.1817 | 0.3029 |
| Postoculars | 0.3175 | 0.3540 |
| Anterior temporal | 0.3243 | 0.0786 |
| Posterior temporal | 0.3693 | 0.2800 |
| Pattern cycles | 0.4324 | –0.0104 |
| Frontal pitting | 0.0859 | –0.4338 |
| Inter-prefrontal sulcus | 0.2073 | 0.2637 |
Table 11. Observed (rows) and predicted (columns) classifications of male Dasypeltis from the Horn of Africa and Egypt according to the Discriminant Function Analysis.
| % correct | D.
atra |
D. scabra | D. taylori | D. abyssina | D. arabica | D.
bazi |
|
| p = .280 | p = .240 | p = .120 | p = .160 | p = .080 | p = .120 | ||
| D. atra | 100 | 7 | 0 | 0 | 0 | 0 | 0 |
| D. scabra | 100 | 0 | 6 | 0 | 0 | 0 | 0 |
| D. taylori | 100 | 0 | 0 | 3 | 0 | 0 | 0 |
| D. abyssina | 100 | 0 | 0 | 0 | 4 | 0 | 0 |
| D. arabica | 100 | 0 | 0 | 0 | 0 | 2 | 0 |
| D. bazi | 66.7 | 1 | 0 | 0 | 0 | 0 | 2 |
| Total | 96.0 | 8 | 6 | 3 | 4 | 2 | 2 |
Table 12. Standardised Canonical Discriminant Function Coefficients (Sigma-restricted parameterisation) for the analysis of male Dasypeltis in the Horn of Africa and Egypt.
| Function 1 | Function 2 | |
| Midbody scale rows | 2.08508 | –0.334990 |
| Ventrals | 2.51335 | –0.269100 |
| Subcaudals | 0.08589 | –0.155450 |
| Supralabials | 0.05782 | –0.447450 |
| Postoculars | –1.36866 | 0.590286 |
| Anterior temporal | 1.34341 | 0.105930 |
| Posterior temporal | 0.11259 | 0.006628 |
| Pattern cycles | –0.70219 | –0.824390 |
| Frontal pitting | 0.13149 | –0.68330003 |
| Inter-prefrontal sulcus | –0.36369 | –0.534840 |
| Eigenvalue | 42.25546 | 2.395576 |
| Cumulative Proportion | 0.88362 | 0.933713 |
Table 13. Observed (rows) and predicted (columns) classifications of female Dasypeltis from the Horn of Africa and Egypt according to the Discriminant Function Analysis.
| % correct | D.
atra |
D. scabra | D. taylori | D. abyssina | D. arabica | D.
bazi |
D. crucifera | |
| p = .237 | p = .316 | p = .053 | p = .184 | p = .105 | p = .053 | p = .053 | ||
| D. atra | 77.8 | 7 | 1 | 0 | 1 | 0 | 0 | 0 |
| D. scabra | 75.0 | 1 | 9 | 0 | 0 | 1 | 1 | 0 |
| D. taylori | 100 | 0 | 0 | 2 | 0 | 0 | 0 | 0 |
| D. abyssina | 100 | 0 | 0 | 0 | 7 | 0 | 0 | 0 |
| D. arabica | 75.0 | 0 | 1 | 0 | 0 | 3 | 0 | 0 |
| D. bazi | 50.0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 |
| D. crucifera | 100 | 0 | 0 | 0 | 0 | 0 | 0 | 2 |
| Total | 81.6 | 8 | 12 | 2 | 8 | 4 | 2 | 2 |
Table 14. Standardised Canonical Discriminant Function Coefficients (Sigma-restricted parameterisation) for the analysis of female Dasypeltis in the Horn of Africa and Egypt.
| Function 1 | Function 2 | |
| Midbody scale rows | –0.187479 | 0.117839 |
| Ventrals | 1.074553 | 0.262493 |
| Subcaudals | –0.216661 | 1.018205 |
| Supralabials | –0.492329 | –0.208635 |
| Preoculars | 0.097186 | 0.420874 |
| Postoculars | 0.178724 | –0.409874 |
| Anterior temporal | –0.063169 | 0.143802 |
| Posterior temporal | 0.379009 | 0.083376 |
| Pattern cycles | –0.460762 | 0.680089 |
| Frontal pitting | –0.298385 | –0.084354 |
| Inter-prefrontal sulcus | –0.280103 | 0.377127 |
| Eigenvalue | 3.706452 | 2.549360 |
| Cumulative Proportion | 0.462892 | 0.781277 |
Appendix 1. Morphological data for specimens of Dasypeltis used in this study. Museum catalogue numbers with asterisks denote specimens with full sets of data used in the multivariate analyses; a few specimens with full data, obtained subsequent to these analyses, are marked by +. Under ‘Sex’: M = male, F = female, U = undetermined. Counts for supralabials, preoculars, postoculars, anterior temporals and posterior temporals include both sides of the head. Under ‘Cycles’: B denotes plain brown or greyish specimens with no discernible dorsal pattern; for D. medici, patterned specimens for which cycle counts are not available are indicated as P, while dorsally uniform or weakly marked specimens are coded as L. For D. atra, patterned specimens for which cycle counts are not available are indicated as ‘2Bx’, while specimens of the ‘lineolata’ morph (a variety of ‘2Bx’) are indicated as ‘lin’ (see Gans 1959). Frontal pitting: 0 = smooth, 1 = marginally pitted (States 1 & 2), 2 = extensively pitted (States 3 & 4); Inter-prefrontal sulcus: 0 = weakly marked, 1 = moderately marked, 2 = deeply sunken.
| Species | Catalogue No. | Country | Locality | Sex | SVL | Tail | SVL/Tail | Midbody scale rows | Ventrals | Subcaudals | Supralabials | Preoculars | Postoculars | AnterTemps | PosterTemps | Pattern cycles | Frontal pitting | Inter-prefrontal sulcus |
| D. abyssina | NMW 15301/2* | ERITREA | ? | M | 412 | 74 | 5.568 | 23 | 239 | 60 | 14 | 2 | 4 | 4 | 5 | 77 | 1 | 1 |
| D. abyssina | NMW 15301/3 | ERITREA | ? | M | 504 | 24 | 247 | 14 | 2 | 4 | 4 | 6 | 93 | 1 | 0 | |||
| D. abyssina | BM 69.11.4.38* | ERITREA | Senafe | M | 270 | 45 | 6 | 24 | 239 | 61 | 14 | 2 | 4 | 4 | 5 | 67 | 1 | 0 |
| D. abyssina | FMNH 12728* | ETHIOPIA | Gondar | M | 348 | 57 | 6.105 | 23 | 236 | 57 | 14 | 2 | 4 | 4 | 5 | 95 | 1 | 0 |
| D. abyssina | BM 1902.12.13.85 | ETHIOPIA | Zegie, Lake Tsana | M | 23 | 226 | 2 | 4 | 4 | 8 | 65 | 0 | 0 | |||||
| D. abyssina | BM 1902.12.13.86* | ETHIOPIA | Zegie, Lake Tsana | M | 235 | 44 | 5.341 | 23 | 226 | 68 | 14 | 2 | 2 | 4 | 3 | B | 1 | 0 |
| D. abyssina | NMW 15301/1* | ERITREA | ? | F | 539 | 80 | 6.738 | 23 | 251 | 54 | 14 | 2 | 4 | 4 | 7 | 73 | 1 | 0 |
| D. abyssina | NMW 15300* | ERITREA | Gheleb | F | 540 | 72 | 7.5 | 24 | 271 | 54 | 14 | 2 | 4 | 4 | 7 | 81 | 1 | 0 |
| D. abyssina | MNHN 6567 | ETHIOPIA | ? | F | 665 | 90 | 7.389 | 25 | 260 | |||||||||
| D. abyssina | MSNM 2699A | ETHIOPIA | Gondar | F | 655 | 91 | 7.198 | 22 | 248 | 59 | 14 | 2 | 4 | 4 | B | |||
| D. abyssina | MZUF 783* | ETHIOPIA | Gondar | F | 416 | 52 | 8 | 25 | 254 | 49 | 14 | 2 | 4 | 4 | 6 | 98 | 1 | 1 |
| D. abyssina | NMBO 10256* | ETHIOPIA | Keriyo | F | 422 | 52 | 8.115 | 25 | 264 | 51 | 14 | 2 | 4 | 4 | 6 | 89 | 1 | 0 |
| D. abyssina | NMBO 10257* | ETHIOPIA | Keriyo | F | 483 | 87 | 5.552 | 22 | 241 | 65 | 14 | 2 | 4 | 4 | 7 | B | 1 | 0 |
| D. abyssina | NMZB 17099* | ETHIOPIA | Keriyo | F | 475 | 70 | 6.786 | 22 | 247 | 59 | 14 | 2 | 4 | 3 | 6 | B | 1 | 0 |
| D. abyssina | MSNM 2700A | ETHIOPIA | Mai Canetta | F | 663 | 84 | 7.893 | 21 | 263 | 60 | 14 | 2 | 4 | 4 | B | |||
| D. abyssina | MCZ 80984* | ETHIOPIA | Ras Dashen Terara | F | 511 | 74 | 6.905 | 24 | 261 | 59 | 14 | 2 | 4 | 4 | 6 | B | 1 | 0 |
| D. abyssina | MSNM 2699B | ETHIOPIA | Gondar | U | 364 | 59 | 6.169 | 22 | 222 | 61 | 14 | 2 | 4 | 4 | 73 | |||
| D. abyssina | MSNM 2699C | ETHIOPIA | Gondar | U | 240 | 22 | 228 | 14 | 2 | 4 | 4 | 77 | ||||||
| D. abyssina | MSNM 2700B | ETHIOPIA | Mai Canetta | U | 220 | 28 | 7.857 | 21 | 257 | 58 | 14 | 2 | 4 | 4 | B | |||
| D. arabica | MHNG 2536.42* | SAUDI ARABIA | vicinity of Abha | M | 24 | 238 | 63 | 14 | 2 | 4 | 4 | 6 | 69 | 1 | 1 | |||
| D. arabica | BM 03.6.26.41* | YEMEN | El Kubar | M | 325 | 56 | 5.804 | 23 | 244 | 65 | 14 | 2 | 4 | 4 | 8 | 72 | 1 | 1 |
| D. arabica | MSNG 52221 | YEMEN | El Siyani | M | 486 | 77 | 6.312 | 23 | 236 | 63 | 14 | 2 | 4 | 4 | 6 | |||
| D. arabica | BM 1987.1018* | SAUDI ARABIA | Ajibah | F | 246 | 60 | 4.1 | 23 | 246 | 55 | 14 | 2 | 2 | 4 | 6 | 79 | 1 | 1 |
| D. arabica | MHNG 2536.41* | SAUDI ARABIA | Aqabat Raydah | F | 43 | 25 | 254 | 61 | 14 | 2 | 4 | 4 | 6 | 71 | 1 | 2 | ||
| D. arabica | BM 1987.2192* | YEMEN | Sana’a | F | 625 | 89 | 7.022 | 25 | 240 | 53 | 14 | 4 | 4 | 5 | 7 | 67 | 1 | 2 |
| D. arabica | BM 1987.2193* | YEMEN | Taiz | F | 400 | 64 | 6.25 | 25 | 239 | 58 | 14 | 2 | 4 | 4 | 7 | 62 | 1 | 1 |
| D. atra | MRAC 18583 | BURUNDI | Kamagaba Marsh | M | 457 | 86 | 5.314 | 24 | 208 | 57 | 14 | 2 | 2 | 4 | 5 | 2Bx | 1 | 0 |
| D. atra | MRAC 18585 | BURUNDI | Rumonge | M | 457 | 24 | 215 | 12 | 2 | 2 | 4 | 6 | 2Bx | 1 | 0 | |||
| D. atra | NMW 9964/2+ | ETHIOPIA | ‘Abessinien’ | M | 350 | 65 | 5.385 | 24 | 209 | 57 | 14 | 2 | 4 | 4 | 7 | 76 | 1 | 0 |
| D. atra | MSNG 30412 | ETHIOPIA | Addis Ababa | M | 227 | 50 | 14 | 2 | 4 | 2 | 4 | 0 | 2 | |||||
| D. atra | ZFMK 57537* | ETHIOPIA | Bedele, Ullubabor | M | 505 | 88 | 5.739 | 22 | 220 | 57 | 14 | 2 | 2 | 4 | 7 | M | 1 | 0 |
| D. atra | MNHN 05-209 | ETHIOPIA | Bourka | M | 478 | 23 | 230 | 14 | 2 | 4 | 4 | 6 | B | 1 | 0 | |||
| D. atra | MNHN 05-210* | ETHIOPIA | Bourka | M | 293 | 45 | 6.511 | 22 | 231 | 58 | 14 | 2 | 4 | 4 | 4 | M | 1 | 0 |
| D. atra | BM 1964.1068* | ETHIOPIA | Gemu Gofa District | M | 435 | 75 | 5.8 | 25 | 224 | 58 | 14 | 2 | 4 | 4 | 6 | 85 | 0 | 0 |
| D. atra | ZFMK 16297* | ETHIOPIA | Genale to Chamo | M | 504 | 87 | 5.793 | 25 | 214 | 55 | 14 | 2 | 4 | 4 | 6 | B | 0 | 0 |
| D. atra | BM 1973.3175 | ETHIOPIA | Ghimbi, Wollega | M | 390 | 76 | 5.132 | 61 | 14 | 2 | 4 | 4 | 6 | M | 1 | 0 | ||
| D. atra | MCZ 34923 | ETHIOPIA | Harar | M | 259 | 46 | 5.63 | 23 | 225 | 60 | 14 | 2 | 4 | 4 | 6 | 2Bx | 1 | 1 |
| D. atra | NMW 9964-1* | ETHIOPIA | Harar | M | 352 | 58 | 6.069 | 22 | 232 | 58 | 14 | 2 | 4 | 4 | 6 | B | 1 | 1 |
| D. atra | MNHN 05-205 | ETHIOPIA | Laga Hardine | M | 328 | 58 | 5.656 | 25 | 211 | 61 | 14 | 2Bx | 1 | 0 | ||||
| D. atra | MNHN 05-206* | ETHIOPIA | Laga Hardine | M | 325 | 60 | 5.417 | 25 | 219 | 58 | 14 | 2 | 4 | 4 | 6 | 74 | 1 | 0 |
| D. atra | ZMB 27463 | ETHIOPIA | Lake Alemaya | M | 245 | 40 | 6.125 | 23 | 231 | 60 | 14 | 2 | 4 | 4 | 5 | 2Bx | 1 | 1 |
| D. atra | ZFMK 16237* | ETHIOPIA | Lake Shala | M | 414 | 72 | 5.75 | 23 | 220 | 58 | 14 | 2 | 4 | 6 | 8 | 79 | 0 | 0 |
| D. atra | MSNG 29047A | ETHIOPIA | Shoa | M | 22 | 216 | 54 | 14 | 2 | 2 | 4 | 1 | 1 | |||||
| D. atra | BM 96.5.19.57 | ETHIOPIA | Shoa (Ragazzi) | M | 315 | 49 | 6.429 | 22 | 221 | 59 | 14 | 2 | 4 | 4 | 7 | M | 0 | |
| D. atra | NMK 483 | KENYA | ? | M | 453 | 96 | 4.719 | 216 | 67 | 14 | 2 | B | ||||||
| D. atra | CAS 148035* | KENYA | Chemelil, Kisumu | M | 420 | 83 | 5.06 | 23 | 209 | 62 | 14 | 2 | 3 | 4 | 6 | M | 1 | 0 |
| D. atra | CAS 122281* | KENYA | Chuka, Meru | M | 505 | 110 | 4.591 | 23 | 201 | 67 | 13 | 2 | 4 | 4 | 6 | B | 1 | 0 |
| D. atra | BM 04.12.23.10* | KENYA | Fort Hall (= Murang’a) | M | 183 | 33 | 5.545 | 26 | 221 | 67 | 14 | 2 | 4 | 4 | 6 | 106 | 1 | 0 |
| D. atra | NMK 3329 | KENYA | Kabarnet, Baringo | M | 454 | 199 | 14 | 2 | B | |||||||||
| D. atra | NMK 479 | KENYA | Kabete, Kiambu | M | 520 | 97 | 5.361 | 215 | 57 | 14 | 2 | B | ||||||
| D. atra | MCZ 40577* | KENYA | Kaimosi | M | 346 | 64 | 5.406 | 24 | 211 | 65 | 14 | 2 | 3 | 4 | 6 | M | 1 | 0 |
| D. atra | CAS 142248* | KENYA | Kakamega Forest Stn. | M | 515 | 97 | 5.309 | 23 | 209 | 59 | 14 | 2 | 4 | 4 | 6 | M | 1 | 0 |
| D. atra | NMK 3007 | KENYA | Karen, Nairobi | M | 464 | 93 | 4.989 | 206 | 61 | 14 | 4 | B | ||||||
| D. atra | NMK 2952/1 | KENYA | Kiboko Ranch Station | M | 198 | 37 | 5.351 | 206 | 67 | 14 | 2 | 2Bx | ||||||
| D. atra | NMK 2862 | KENYA | Kiganjo, Nyeri | M | 428 | 81 | 5.284 | 200 | 55 | 14 | 2 | M | ||||||
| D. atra | CG 2278 | KENYA | Lumbwa, Pen-y-Bryn | M | 508 | 99 | 5.131 | 23 | 213 | 62 | 2 | 3 | 4 | 6 | 106 | 1 | 0 | |
| D. atra | CG 2279 | KENYA | Lumbwa, Pen-y-Bryn | M | 285 | 47 | 6.064 | 24 | 213 | 58 | 2 | 4 | 6 | 7 | 62 | 1 | 0 | |
| D. atra | NMK 659 | KENYA | Mount Elgon | M | 434 | 94 | 4.617 | 211 | 60 | B | ||||||||
| D. atra | LACM 50631* | KENYA | Mweiga | M | 476 | 96 | 4.958 | 23 | 210 | 59 | 15 | 2 | 2 | 4 | 6 | B | 1 | 0 |
| D. atra | NMK 3339 | KENYA | Nairobi | M | 483 | 90 | 5.367 | 222 | 61 | 14 | 2 | 76 | ||||||
| D. atra | NMK 3548 | KENYA | Nairobi | M | 421 | 85 | 4.953 | 212 | 62 | 14 | 2 | 95 | ||||||
| D. atra | NMK 403 | KENYA | Nairobi | M | 384 | 66 | 5.818 | 207 | 59 | 14 | 2 | B | ||||||
| D. atra | BM 1960.1.6.40* | KENYA | Ngong | M | 420 | 83 | 5.06 | 23 | 209 | 65 | 14 | 2 | 4 | 4 | 6 | 88 | 1 | 1 |
| D. atra | NMK 2576 | KENYA | North Nandi Forest, Chemisia | M | 456 | 98 | 4.653 | 208 | 59 | 14 | 2 | M | ||||||
| D. atra | NMK 2590 | KENYA | North Nandi Forest, Chemisia | M | 466 | 99 | 4.707 | 208 | 60 | 14 | 2 | |||||||
| D. atra | NMK 1497 | KENYA | Nyambene Hills, Meru | M | 421 | 87 | 4.839 | 207 | 61 | 12 | 2 | B | ||||||
| D. atra | NMK 4506 | KENYA | Ol Ari Nyiro Ranch | M | 365 | 26 | 210 | 14 | 2 | 4 | 4 | 6 | 74 | 1 | 0 | |||
| D. atra | NMK 4176 | KENYA | South Nandi Forest | M | 465 | 71 | 6.549 | 208 | 50 | 14 | 2 | M | ||||||
| D. atra | FMNH 58337 | KENYA | Subukia | M | 546 | 110 | 4.964 | 23 | 212 | 60 | 14 | 2 | 2 | 4 | 4 | 2Bx | 1 | |
| D. atra | MRAC 18554 | RWANDA | Astrida | M | 297 | 54 | 5.5 | 23 | 210 | 61 | 14 | 2 | 2 | 4 | 6 | 2Bx | 1 | 0 |
| D. atra | PEM R5623 | RWANDA | Btwn Gikongoro & Nyungwe For. R. | M | 429 | 81 | 5.296 | 23 | 211 | 59 | 14 | 3 | 3 | 4 | 6 | 2Bx | 1 | 0 |
| D. atra | ZFMK 61616* | RWANDA | Bugesera | M | 426 | 84 | 5.071 | 24 | 216 | 62 | 14 | 2 | 2 | 4 | 6 | M | 1 | 0 |
| D. atra | MRAC 2723 | RWANDA | Kisenyi | M | 70 | 22 | 207 | 66 | 14 | 3 | 2 | 4 | 5 | 2Bx | 1 | 0 | ||
| D. atra | MRAC 2725 | RWANDA | Kisenyi | M | 404 | 96 | 4.208 | 22 | 207 | 66 | 14 | 2 | 2 | 2 | 4 | 2Bx | 1 | 0 |
| D. atra | MRAC 2727* | RWANDA | Kisenyi | M | 414 | 88 | 4.705 | 23 | 218 | 63 | 13 | 2 | 2 | 4 | 5 | B | 1 | 0 |
| D. atra | MRAC 3821* | RWANDA | Kisenyi | M | 383 | 83 | 4.614 | 24 | 210 | 66 | 14 | 2 | 2 | 4 | 4 | B | 1 | 0 |
| D. atra | MRAC 5871 | RWANDA | Kisenyi | M | 413 | 94 | 4.394 | 24 | 208 | 66 | 14 | 2 | 2 | 4 | 7 | 2Bx | 1 | 0 |
| D. atra | MRAC 15477 | RWANDA | Nyanza | M | 226 | 40 | 5.65 | 24 | 212 | 63 | 14 | 2 | 3 | 5 | 6 | 2Bx | 1 | 0 |
| D. atra | MRAC 14331 | RWANDA | Rwankeri District | M | 352 | 61 | 5.77 | 23 | 212 | 60 | 14 | 2 | 2 | 4 | 6 | 2Bx | 1 | 0 |
| D. atra | MRAC 18562 | RWANDA | Tare, Astrida | M | 24 | 2 | 4 | 5 | B | 0 | ||||||||
| D. atra | BM 1978.380 | SOUTH SUDAN | Gilo, Imatong Mtns | M | 530 | 104 | 5.1 | 23 | 14 | 2 | 2 | 4 | 4 | M | 1 | 0 | ||
| D. atra | FMNH 62215 | SOUTH SUDAN | Gilo, Imatong Mtns | M | 501 | 102 | 4.912 | 23 | 219 | 63 | 14 | 2 | 3 | 4 | 6 | M | 1 | |
| D. atra | MCZ 53406* | SOUTH SUDAN | Gilo, Imatong Mtns | M | 536 | 116 | 4.621 | 24 | 210 | 66 | 14 | 2 | 4 | 4 | 6 | M | 1 | 0 |
| D. atra | MCZ 53407 | SOUTH SUDAN | Gilo, Imatong Mtns | M | 459 | 94 | 4.883 | 23 | 212 | 60 | 14 | M | 1 | 0 | ||||
| D. atra | MCZ 53408* | SOUTH SUDAN | Gilo, Imatong Mtns | M | 518 | 116 | 4.466 | 23 | 217 | 65 | 14 | 2 | 2 | 4 | 6 | M | 1 | 0 |
| D. atra | ZFMK 29564 | SOUTH SUDAN | Gilo, Imatong Mtns | M | 24 | 221 | 56 | 15 | 2 | 2 | 4 | 8 | M | |||||
| D. atra | ZFMK 29719 | SOUTH SUDAN | Gilo, Imatong Mtns | M | 24 | 218 | 63 | 14 | 2 | 2 | 4 | 6 | M | |||||
| D. atra | ZMUC 6116 | SOUTH SUDAN | Gilo, Imatong Mtns | M | 493 | 23 | 210 | 14 | 2 | 2 | 4 | 4 | M | 1 | 0 | |||
| D. atra | ZFMK 26005 | SOUTH SUDAN | Katire | M | 22 | 219 | 65 | 14 | 2 | 2 | 4 | 6 | M | 1 | ||||
| D. atra | ZFMK 29731 | SOUTH SUDAN | Talanga Forest | M | 23 | 214 | 66 | 12 | 2 | 2 | 4 | 4 | M | |||||
| D. atra | ZMB 16749* | TANZANIA | Bukoba | M | 398 | 79 | 5.038 | 23 | 213 | 67 | 14 | 2 | 2 | 4 | 5 | 100 | 1 | 0 |
| D. atra | ZMB 16750 | TANZANIA | Bukoba | M | 278 | 48 | 5.792 | 23 | 214 | 59 | 14 | 4 | 2 | 4 | 6 | 2Bx | 1 | 1 |
| D. atra | BM 1994.403* | TANZANIA | Ibaya Camp, Mkomazi G.R. | M | 453 | 82 | 5.524 | 26 | 224 | 59 | 14 | 2 | 4 | 4 | 6 | 90 | 1 | 0 |
| D. atra | NMBO 7375 | TANZANIA | Ibaya Camp, Mkomazi G.R. | M | 211 | 38 | 5.553 | 26 | 220 | 65 | 4 | 4 | 6 | 92 | 1 | 1 | ||
| D. atra | BM 1970.2177 | TANZANIA | Kasoge, Mahale Peninsula | M | 405 | 91 | 4.451 | 25 | 14 | 2 | 2 | 2 | 7 | 109 | 1 | 0 | ||
| D. atra | ZMUC 6122 | TANZANIA | Mbizi Forest Reserve | M | 398 | 23 | 200 | 67 | 14 | 2 | 3 | 4 | 6 | B | 2 | 1 | ||
| D. atra | ZMUC 6139* | TANZANIA | Minziro Forest Reserve | M | 455 | 91 | 5 | 23 | 211 | 66 | 14 | 2 | 2 | 4 | 6 | M | 1 | 0 |
| D. atra | ZSM 81/1936a | TANZANIA | Moshi | M | 355 | 23 | 208 | 14 | 2Bx | |||||||||
| D. atra | ZSM 81/1936b | TANZANIA | Moshi | M | 370 | 75 | 4.933 | 23 | 206 | 58 | 14 | 2Bx | ||||||
| D. atra | LIV 1980.150.30 | UGANDA | ? | M | 445 | 85 | 5.235 | 205 | 58 | 2 | 4 | 4 | B | |||||
| D. atra | LIV 1960.67.1 | UGANDA | Bihunga, Bukwezi | M | 440 | 96 | 4.583 | 215 | 69 | 2 | 6 | 6 | M | |||||
| D. atra | USNM 63481 | UGANDA | Budongo Forest | M | 560 | 115 | 4.87 | 23 | 221 | 63 | 14 | 2 | 3 | 4 | 5 | M | 1 | |
| D. atra | LIV 1962.50.32 | UGANDA | Buhunga, Buweju | M | 494 | 98 | 5.041 | 62 | 2Bx | |||||||||
| D. atra | BM 11.7.8.14* | UGANDA | Bussu, Jinja | M | 480 | 95 | 5.053 | 24 | 212 | 63 | 14 | 2 | 4 | 4 | 6 | M | 1 | 1 |
| D. atra | MSNG 30413B* | UGANDA | Bussu, Jinja | M | 611 | 98 | 6.235 | 23 | 209 | 65 | 14 | 2 | 4 | 4 | 4 | B | 1 | 0 |
| D. atra | BM 51.1.3.74* | UGANDA | Entebbe | M | 183 | 35 | 5.229 | 25 | 202 | 62 | 12 | 2 | 4 | 4 | 6 | B | 1 | 0 |
| D. atra | CAS-SU 21710* | UGANDA | Fort Portal, Kingami | M | 438 | 92 | 4.761 | 23 | 212 | 63 | 14 | 2 | 3 | 4 | 6 | B | 1 | 0 |
| D. atra | LIV 1962.9.28 | UGANDA | Kagabagara, Isingiro | M | 508 | 100 | 5.08 | 210 | 62 | 2 | 4 | 5 | B | |||||
| D. atra | BM 36.7.3.32 | UGANDA | Kampala | M | 463 | 101 | 4.584 | 23 | 208 | 64 | 14 | 2 | 2 | 4 | 6 | 2Bx | 1 | 0 |
| D. atra | LIV 1980.150.31 | UGANDA | Kayonza, Kinkizi, Kigezi | M | 292 | 53 | 5.509 | 208 | 61 | 2 | 4 | 6 | B | |||||
| D. atra | NMZB-UM 2550* | UGANDA | Kayonza, Kinkizi, Kigezi | M | 450 | 92 | 4.891 | 22 | 210 | 63 | 14 | 2 | 3 | 4 | 6 | M | 1 | 0 |
| D. atra | NMK 404 | UGANDA | Kazinga Channel | M | 380 | 87 | 4.368 | 221 | 66 | 14 | 2 | B | ||||||
| D. atra | LIV 1962.9.14 | UGANDA | Kichwamba, Bunyaruguru | M | 348 | 60 | 5.8 | 210 | 57 | 4 | 4 | 6 | 2Bx | |||||
| D. atra | ZMB 27539 | UGANDA | Kisenyi | M | 224 | 43 | 5.209 | 23 | 217 | 67 | 14 | 2 | 2 | 4 | 6 | 2Bx | 1 | 0 |
| D. atra | BM 1961.1805* | UGANDA | Kyamuhunga, Igara, Ankole | M | 405 | 83 | 4.88 | 23 | 208 | 63 | 14 | 2 | 2 | 4 | 5 | M | 1 | 0 |
| D. atra | FMNH 9914 | UGANDA | Lake Bunyonyi | M | 446 | 86 | 5.186 | 24 | 211 | 61 | 14 | 2 | 2 | 4 | 8 | 2Bx | 1 | |
| D. atra | BM 32.5.2.99* | UGANDA | Lake George | M | 450 | 104 | 4.327 | 24 | 208 | 71 | 14 | 2 | 3 | 4 | 6 | B | 1 | 0 |
| D. atra | BM 51.1.3.78* | UGANDA | Lake Nabugabo | M | 468 | 92 | 5.087 | 23 | 220 | 69 | 14 | 2 | 2 | 4 | 5 | B | 1 | 0 |
| D. atra | LIV 1980.150.27 | UGANDA | Lukiri, Mitoma | M | 447 | 85 | 5.259 | 215 | 61 | 2 | 4 | 6 | M | |||||
| D. atra | BM 08.10.20.14* | UGANDA | Mabira Forest, Chagwe | M | 179 | 30 | 5.967 | 25 | 218 | 61 | 14 | 2 | 2 | 4 | 6 | B | 1 | 0 |
| D. atra | BM 03.12.2.23* | UGANDA | Msozi – Isingiro | M | 455 | 98 | 4.643 | 24 | 214 | 67 | 14 | 2 | 2 | 3 | 5 | M | 1 | 0 |
| D. atra | BM 34.12.15.585* | UGANDA | Muko, Kigezi | M | 355 | 65 | 5.462 | 23 | 205 | 61 | 14 | 2 | 2 | 4 | 6 | M | 0 | 0 |
| D. atra | BM 34.12.15.586* | UGANDA | Muko, Kigezi | M | 410 | 82 | 5 | 24 | 208 | 64 | 14 | 4 | 2 | 4 | 4 | 85 | 1 | 0 |
| D. atra | BM 34.12.15.587* | UGANDA | Muko, Kigezi | M | 201 | 38 | 5.289 | 23 | 206 | 65 | 14 | 2 | 2 | 4 | 6 | 107 | 1 | 0 |
| D. atra | BM 34.12.15.589 | UGANDA | Muko, Kigezi | M | 215 | 43 | 5 | 24 | 204 | 68 | 14 | 2 | 2 | 4 | 5 | 2Bx | 1 | 0 |
| D. atra | BM 34.12.15.590 | UGANDA | Muko, Kigezi | M | 420 | 23 | 211 | 14 | 2 | 3 | 4 | 8 | M | 1 | 0 | |||
| D. atra | BM 34.12.15.591* | UGANDA | Muko, Kigezi | M | 455 | 88 | 5.17 | 23 | 207 | 64 | 14 | 2 | 2 | 4 | 5 | B | 1 | 0 |
| D. atra | BM 34.12.15.593 | UGANDA | Muko, Kigezi | M | 327 | 59 | 5.542 | 23 | 218 | 63 | 14 | 2 | 2 | 4 | 7 | 2Bx | 0 | |
| D. atra | MCZ 48363 | UGANDA | Mushongero, Lake Mutanda | M | 320 | 63 | 5.079 | 24 | 206 | 63 | 14 | 2 | 2 | 4 | 6 | 2Bx | 1 | 0 |
| D. atra | LIV 1980.150.26 | UGANDA | Nsika, Buhwezi | M | 320 | 212 | 3 | 4 | 6 | M | ||||||||
| D. atra | LIV 1962.9.23 | UGANDA | Nyabushozi | M | 400 | 83 | 4.819 | 213 | 65 | 2 | 4 | 6 | M | |||||
| D. atra | UMMZ 88532/1 | UGANDA | Nyakabende | M | 236 | 35 | 6.743 | 24 | 205 | 63 | 14 | 2 | 2 | 4 | 4 | 2Bx | 1 | |
| D. atra | LIV 1962.50.33 | UGANDA | Nyansimbo, Mitoma | M | 328 | 62 | 5.29 | 206 | 62 | 4 | 4 | 5 | B | |||||
| D. atra | NMZB-UM 2549 | UGANDA | Nyanuhunga, Igara, Ankole | M | 430 | 96 | 4.479 | 24 | 204 | 63 | 14 | 2 | 2 | 4 | 6 | 2Bx | 1 | 0 |
| D. atra | LIV 1980.150.29 | UGANDA | Nyarusiza, Bufumbira, Kigezi | M | 460 | 99 | 4.646 | 220 | 65 | 2 | 4 | 4 | 2Bx | |||||
| D. atra | NMZB-UM 2553* | UGANDA | Nyarusiza, Bufumbira, Kigezi | M | 320 | 58 | 5.517 | 22 | 208 | 62 | 14 | 2 | 2 | 4 | 5 | 103 | 1 | 0 |
| D. atra | MSNG 40764* | UGANDA | Sesse Islands, Lake Victoria | M | 472 | 77 | 6.13 | 23 | 214 | 67 | 14 | 2 | 4 | 4 | 7 | B | 1 | 0 |
| D. atra | MCZ 40567* | UGANDA | Sipi, Mount Elgon | M | 212 | 39 | 5.436 | 23 | 208 | 62 | 14 | 2 | 2 | 4 | 4 | B | 1 | 0 |
| D. atra | MCZ 40568 | UGANDA | Sipi, Mount Elgon | M | 473 | 90 | 5.256 | 23 | 219 | 64 | 14 | 2 | 2 | 4 | 6 | 2Bx | 0 | |
| D. atra | MCZ 40570* | UGANDA | Sipi, Mount Elgon | M | 518 | 97 | 5.34 | 23 | 211 | 59 | 14 | 2 | 3 | 4 | 4 | B | 1 | 0 |
| D. atra | MRAC 82-16-R-3* | BURUNDI | Bujumbura | F | 615 | 100 | 6.15 | 25 | 215 | 57 | 14 | 2 | 2 | 4 | 6 | 130 | 0 | 0 |
| D. atra | MRAC 18584* | BURUNDI | Makamba | F | 184 | 30 | 6.133 | 24 | 219 | 56 | 14 | 2 | 2 | 4 | 6 | B | 1 | 0 |
| D. atra | BM 16.6.24.13* | ETHIOPIA | Aware Melca | F | 247 | 31 | 7.968 | 26 | 239 | 52 | 14 | 2 | 2 | 4 | 7 | B | 1 | 0 |
| D. atra | MNHN 05-208 | ETHIOPIA | Bourka | F | 712 | 96 | 7.417 | 22 | 243 | 52 | 13 | 2 | 3 | 3 | 4 | 2Bx | 1 | 0 |
| D. atra | MSNM 2702 | ETHIOPIA | Circao, Dambria | F | 530 | 22 | 232 | 14 | 2 | 4 | 4 | 10 | B | |||||
| D. atra | AMNH 20345* | ETHIOPIA | Ganame | F | 660 | 93 | 7.097 | 22 | 256 | 56 | 15 | 2 | 4 | 4 | 6 | M | 1 | 0 |
| D. atra | NMW 26879/2+ | ETHIOPIA | Ginir | F | 346 | 46 | 7.522 | 26 | 231 | 50 | 13 | 2 | 4 | 4 | 7 | 82 | 1 | 1 |
| D. atra | ZFMK 5535* | ETHIOPIA | Gololtscha | F | 666 | 83 | 8.024 | 23 | 250 | 50 | 15 | 2 | 4 | 4 | 6 | M | 1 | 1 |
| D. atra | NMW 26879/3 | ETHIOPIA | Gumaydo | F | 609 | 28 | 234 | 14 | 2 | 4 | 4 | 4 | 81 | 1 | 0 | |||
| D. atra | FMNH 4012* | ETHIOPIA | Harar | F | 248 | 35 | 7.086 | 26 | 231 | 53 | 14 | 2 | 4 | 4 | 5 | 85 | 1 | 0 |
| D. atra | ZFMK 84993* | ETHIOPIA | Jigjiga | F | 490 | 64 | 7.656 | 27 | 222 | 45 | 14 | 2 | 4 | 4 | 8 | 72 | 1 | 0 |
| D. atra | MNHN 05-211 | ETHIOPIA | Kounni | F | 178 | 24 | 7.417 | 27 | 219 | 48 | 14 | 2 | 4 | 4 | 4 | 2Bx | 1 | 0 |
| D. atra | MNHN 05-207* | ETHIOPIA | Laga Hardine | F | 264 | 34 | 7.765 | 26 | 228 | 51 | 14 | 2 | 4 | 4 | 6 | B | 1 | 0 |
| D. atra | ZFMK 16236* | ETHIOPIA | Lake Abiata | F | 532 | 67 | 7.94 | 25 | 244 | 50 | 14 | 2 | 4 | 4 | 8 | 72 | 1 | 0 |
| D. atra | ZMB 10286* | ETHIOPIA | Let Marefia | F | 266 | 32 | 8.313 | 22 | 243 | 48 | 14 | 2 | 2 | 4 | 4 | M | 1 | 0 |
| D. atra | NMW 15299* | ETHIOPIA | Wondo | F | 708 | 85 | 8.329 | 23 | 239 | 47 | 14 | 2 | 2 | 4 | 4 | M | 2 | 0 |
| D. atra | UMMZ 96274 | KENYA | ? | F | 610 | 93 | 6.559 | 24 | 221 | 55 | 12 | 2 | 4 | 2 | 7 | M | 1 | |
| D. atra | LACM 66076* | KENYA | 13 km SSW of Maralal | F | 244 | 34 | 7.176 | 25 | 225 | 53 | 14 | 4 | 4 | 4 | 6 | 81 | 1 | 0 |
| D. atra | NMW 9964/20+ | KENYA | Athi Plain | F | 478 | 64 | 7.469 | 27 | 223 | 47 | 14 | 2 | 2 | 4 | 6 | B | 1 | 0 |
| D. atra | NMK 3085 | KENYA | Athi River | F | 416 | 59 | 7.051 | 245 | 54 | 14 | 4 | B | ||||||
| D. atra | NMK 3129 | KENYA | Athi-Kapiti Plains | F | 461 | 71 | 6.493 | 228 | 58 | 14 | 4 | B | ||||||
| D. atra | NMK 4174 | KENYA | Baringo | F | 198 | 32 | 6.188 | 231 | 49 | 14 | 4 | B | ||||||
| D. atra | NMK 2323 | KENYA | Butere, Kakamega | F | 314 | 49 | 6.408 | 234 | 53 | 14 | 4 | 79 | ||||||
| D. atra | CAS 148034 | KENYA | Chemelil, Kisumu | F | 605 | 90 | 6.722 | 25 | 225 | 14 | 2 | 4 | 2 | 4 | M | 1 | 1 | |
| D. atra | NMK 406 | KENYA | Chyulu Hills, Makueni | F | 571 | 82 | 6.963 | 228 | 51 | 14 | 2 | B | ||||||
| D. atra | MCZ 40575 | KENYA | Elgonyi | F | 310 | 43 | 7.209 | 24 | 223 | 51 | 14 | 2 | 4 | 4 | 6 | 2Bx | 1 | 0 |
| D. atra | NMK 2849 | KENYA | Embakasi, Nairobi | F | 232 | 58 | 14 | 4 | B | |||||||||
| D. atra | BM 08.9.17.4 | KENYA | Fort Hall (= Murang’a) | F | 500 | 27 | 230 | 14 | 2 | 4 | 4 | 6 | lin | 1 | 0 | |||
| D. atra | NMK 3131 | KENYA | Hopcroft Ranch, Athi River | F | 512 | 75 | 6.827 | 244 | 57 | 12 | 2 | B | ||||||
| D. atra | USNM 40897 | KENYA | Juja Farm | F | 575 | 72 | 7.986 | 26 | 241 | 55 | 15 | 2 | 4 | 4 | 6 | lin | 1 | |
| D. atra | BM 1957.1.11.42* | KENYA | Kabarnet, Baringo | F | 595 | 92 | 6.467 | 23 | 236 | 56 | 12 | 2 | 2 | 4 | 6 | M | 1 | 0 |
| D. atra | CAS 111767 | KENYA | Kabartonjo, Kamasia Mtns | F | 645 | 102 | 6.324 | 25 | 229 | 55 | 13 | 2 | 2 | 4 | 6 | B | 1 | |
| D. atra | NMK 486 | KENYA | Kacheliba | F | 440 | 71 | 6.197 | 228 | 58 | 14 | 4 | 2Bx | ||||||
| D. atra | MCZ 40576* | KENYA | Kaimosi | F | 700 | 101 | 6.931 | 24 | 220 | 51 | 12 | 2 | 2 | 4 | 6 | M | 1 | 0 |
| D. atra | MCZ 40578* | KENYA | Kaimosi | F | 707 | 98 | 7.214 | 24 | 225 | 55 | 13 | 2 | 4 | 4 | 7 | M | 1 | 0 |
| D. atra | USNM 49078 | KENYA | Kaimosi | F | 724 | 97 | 7.464 | 24 | 217 | 50 | 14 | 2 | 2 | 4 | 4 | M | 1 | |
| D. atra | MCZ 18205 | KENYA | Kaimosi (Yala River) | F | 610 | 24 | 221 | 14 | 2 | 3 | 4 | 4 | 2Bx | 1 | 0 | |||
| D. atra | UMMZ 61224 | KENYA | Kaimosi (Yala River) | F | 620 | 88 | 7.045 | 23 | 223 | 50 | 12 | 2 | 2 | 4 | 6 | M | 1 | |
| D. atra | LACM 93378* | KENYA | Kairuni, Nyambeni Mts | F | 662 | 114 | 5.807 | 26 | 222 | 56 | 14 | 3 | 4 | 4 | 6 | 102 | 1 | 1 |
| D. atra | LACM 93379* | KENYA | Kairuni, Nyambeni Mts | F | 203 | 35 | 5.8 | 26 | 226 | 57 | 14 | 2 | 4 | 4 | 7 | B | 1 | 0 |
| D. atra | BM 1959.1.5.58 | KENYA | Kakamega | F | 665 | 105 | 6.333 | 221 | 51 | 14 | 2 | 4 | 4 | 8 | M | 0 | ||
| D. atra | USNM 49376 | KENYA | Kakamega | F | 448 | 74 | 6.054 | 22 | 213 | 55 | 13 | 2 | 4 | 2 | 4 | M | 1 | |
| D. atra | NMK 2503 | KENYA | Karen, Nairobi | F | 550 | 92 | 5.978 | 223 | 56 | 12 | 2 | B | ||||||
| D. atra | NMK 2952/3 | KENYA | Kiboko Ranch Station | F | 175 | 27 | 6.481 | 216 | 51 | 14 | 2 | 2Bx | ||||||
| D. atra | NMK 2952/4 | KENYA | Kiboko Ranch Station | F | 230 | 247 | 14 | 2 | 2Bx | |||||||||
| D. atra | NMK 2130 | KENYA | Kijabe | F | 584 | 95 | 6.147 | 219 | 51 | 12 | 2 | B | ||||||
| D. atra | ZMUC 6101* | KENYA | Kitale | F | 647 | 91 | 7.11 | 25 | 225 | 51 | 14 | 2 | 2 | 4 | 4 | M | 1 | 0 |
| D. atra | NMK 3413 | KENYA | Kitengela, Athi River | F | 581 | 77 | 7.545 | 232 | 58 | 14 | 2 | B | ||||||
| D. atra | ZMB 9224 | KENYA | Kitui | F | 555 | 67.5 | 8.162 | 27 | 233 | 49 | 14 | 2 | 4 | 4 | 6 | lin | 1 | 0 |
| D. atra | BM 1932.5.2.100* | KENYA | Lake Naivasha | F | 640 | 94 | 6.809 | 230 | 50 | 14 | 2 | 4 | 4 | 5 | M | 1 | 0 | |
| D. atra | BM 93.11.21.72* | KENYA | Lake Naivasha | F | 672 | 93 | 7.226 | 25 | 223 | 52 | 14 | 2 | 4 | 4 | 7 | M | 1 | 0 |
| D. atra | NMK 3109 | KENYA | Lorgorien, Trans-Mara | F | 655 | 101 | 6.485 | 225 | 52 | 14 | 4 | B | ||||||
| D. atra | USNM 62907 | KENYA | Mbunyi | F | 765 | 23 | 227 | 13 | 2 | 2 | 4 | M | 1 | 0 | ||||
| D. atra | NMK 657 | KENYA | Mount Elgon | F | 539 | 85 | 6.341 | 217 | 53 | 14 | 2 | M | ||||||
| D. atra | USNM 40990 | KENYA | Mount Kenya | F | 622 | 95 | 6.547 | 26 | 231 | 54 | 14 | 2 | 4 | 4 | 6 | M | 1 | |
| D. atra | LACM 66395* | KENYA | Mount Kulal | F | 464 | 68 | 6.824 | 24 | 233 | 54 | 14 | 2 | 4 | 4 | 8 | 102 | 1 | 0 |
| D. atra | NMK 1965 | KENYA | Mwino, Sigor | F | 359 | 55 | 6.527 | 235 | 52 | 14 | 2 | B | ||||||
| D. atra | PEM 873* | KENYA | Nairobi | F | 650 | 96 | 6.771 | 23 | 219 | 54 | 13 | 2 | 2 | 4 | 4 | B | 1 | 0 |
| D. atra | MCZ 54108 | KENYA | Nairobi (?) | F | 650 | 86 | 7.558 | 233 | 53 | 14 | 2 | 4 | 4 | 8 | lin | 1 | 0 | |
| D. atra | NMK 2486 | KENYA | Nairobi (Westlands) | F | 524 | 89 | 5.888 | 236 | 54 | 14 | 2 | 105 | ||||||
| D. atra | BM 01-1-3-9 | KENYA | Naitolia (6000 feet) | F | 495 | 27 | 238 | 14 | 2 | 4 | 4 | 7 | 81 | 1 | 0 | |||
| D. atra | NMK 1453 | KENYA | Nakuru | F | 604 | 97 | 6.227 | 218 | 53 | 14 | 2 | B | ||||||
| D. atra | NMK 484 | KENYA | Nakuru | F | 554 | 89 | 6.225 | 220 | 55 | 14 | 4 | B | ||||||
| D. atra | BM 06.7.6.7* | KENYA | Nandi | F | 384 | 53 | 7.245 | 23 | 218 | 50 | 14 | 2 | 2 | 3 | 5 | M | 1 | 0 |
| D. atra | NMK 1158 | KENYA | Nandi Hills | F | 700 | 105 | 6.667 | 224 | 50 | 14 | 4 | M | ||||||
| D. atra | NMK 3490 | KENYA | Nanyuki River, Laikipia | F | 531 | 92 | 5.772 | 224 | 53 | 14 | 2 | M | ||||||
| D. atra | NMK 3512 | KENYA | Nanyuki River, Laikipia | F | 611 | 223 | 14 | 4 | M | |||||||||
| D. atra | NMK 1960 | KENYA | Nbabibi, Naivasha | F | 469 | 58 | 8.086 | 231 | 48 | 14 | 4 | B | ||||||
| D. atra | NMK 2583 | KENYA | North Nandi Forest, Chemisia | F | 890 | 115 | 7.739 | 218 | 53 | 14 | 4 | M | ||||||
| D. atra | CG 2281 | KENYA | Pen-y-Bryn Farm , Lumbwa | F | 705 | 97 | 7.268 | 26 | 231 | 49 | 2 | 4 | 4 | 6 | M | 1 | ||
| D. atra | BM 96.3.27.18* | KENYA | Uganda Railway | F | 232 | 35 | 6.629 | 24 | 212 | 51 | 14 | 2 | 2 | 4 | 5 | 104 | 1 | 0 |
| D. atra | MRAC 18555 | RWANDA | Astrida | F | 613 | 83 | 7.386 | 24 | 224 | 58 | 14 | 2 | 2 | 4 | 4 | 2Bx | 1 | 0 |
| D. atra | MRAC 18556 | RWANDA | Astrida | F | 40 | 25 | 50 | 14 | 2 | 2 | 4 | 6 | 2Bx | 1 | 0 | |||
| D. atra | MRAC 18557 | RWANDA | Astrida | F | 372 | 56 | 6.643 | 24 | 214 | 51 | 12 | 2 | 2 | 4 | 4 | 2Bx | 1 | 1 |
| D. atra | MRAC 18558 | RWANDA | Astrida | F | 207 | 33 | 6.273 | 24 | 222 | 54 | 14 | 2 | 2 | 4 | 5 | 2Bx | 1 | 0 |
| D. atra | MRAC 86-01-R-10 | RWANDA | Bugarama | F | 567 | 90 | 6.3 | 25 | 219 | 54 | 14 | 3 | 2 | 4 | 7 | 2Bx | 1 | 0 |
| D. atra | MRAC 86-01-R-11 | RWANDA | Bugarama | F | 642 | 95 | 6.758 | 24 | 218 | 55 | 14 | 2 | 2 | 4 | 6 | 2Bx | 1 | 0 |
| D. atra | MRAC 18577 | RWANDA | Bulimbi | F | 24 | 221 | 14 | 2 | 2 | 4 | 6 | 2Bx | 1 | 0 | ||||
| D. atra | ZFMK 61617 | RWANDA | Cyamudongo | F | 656 | 100 | 6.56 | 24 | 230 | 58 | 12 | 2 | 2 | 4 | 5 | 2Bx | 1 | 0 |
| D. atra | MRAC 14329 | RWANDA | Kamohorora District | F | 293 | 46 | 6.37 | 24 | 234 | 58 | 14 | 2 | 3 | 4 | 6 | 2Bx | 1 | 0 |
| D. atra | ZFMK 55105* | RWANDA | Kibungo, Rusumo | F | 432 | 68 | 6.353 | 24 | 224 | 55 | 14 | 2 | 2 | 4 | 5 | B | 1 | 1 |
| D. atra | MCZ 48364* | RWANDA | Kisenyi | F | 572 | 91 | 6.286 | 25 | 227 | 59 | 14 | 2 | 2 | 4 | 5 | 115 | 1 | 0 |
| D. atra | MRAC 5872 | RWANDA | Kisenyi | F | 424 | 65 | 6.523 | 24 | 222 | 50 | 14 | 2 | 2 | 4 | 6 | 2Bx | 1 | 0 |
| D. atra | MRAC 9773 | RWANDA | Kisenyi | F | 755 | 115 | 6.565 | 24 | 236 | 53 | 14 | 2 | 2 | 4 | 6 | 2Bx | 1 | 0 |
| D. atra | ZMB 20458 | RWANDA | Kisenyi | F | 517 | 75 | 6.893 | 24 | 222 | 51 | 14 | 2 | 2 | 4 | 5 | 2Bx | 1 | 0 |
| D. atra | ZMB 27156 | RWANDA | Kisenyi | F | 409 | 67 | 6.104 | 25 | 221 | 54 | 14 | 2 | 2 | 4 | 6 | 2Bx | 1 | 0 |
| D. atra | MRAC 14333 | RWANDA | Lake Gando | F | 663 | 103 | 6.437 | 24 | 226 | 54 | 14 | 2 | 2 | 4 | 5 | 2Bx | 1 | 0 |
| D. atra | MRAC 14341 | RWANDA | Lake Gando | F | 65 | 25 | 218 | 54 | 14 | 2 | 2 | 4 | 4 | 2Bx | 1 | 0 | ||
| D. atra | MRAC 7509 | RWANDA | Mutura, Mount Virunga | F | 587 | 90 | 6.522 | 24 | 229 | 59 | 14 | 2 | 2 | 4 | 4 | 2Bx | 1 | 0 |
| D. atra | MRAC 74-23-R-22 | RWANDA | Mwogo near Butare | F | 24 | 217 | 14 | 2 | 2 | 4 | 4 | 2Bx | 1 | 0 | ||||
| D. atra | MRAC 14340 | RWANDA | Rwankeri District | F | 377 | 58 | 6.5 | 24 | 219 | 52 | 14 | 2 | 2 | 4 | 5 | 2Bx | 1 | 0 |
| D. atra | MRAC 15440 | RWANDA | Rwankeri District | F | 673 | 109 | 6.174 | 24 | 225 | 60 | 16 | 2 | 2 | 4 | 7 | 113 | 1 | |
| D. atra | MRAC 18259* | RWANDA | Rwankuba | F | 632 | 100 | 6.32 | 25 | 228 | 54 | 14 | 2 | 2 | 4 | 7 | B | 1 | 0 |
| D. atra | MRAC 4074* | RWANDA | Shangugu | F | 525 | 85 | 6.176 | 24 | 225 | 59 | 14 | 2 | 2 | 4 | 8 | 93 | 1 | 0 |
| D. atra | MRAC 4075* | RWANDA | Shangugu | F | 373 | 58 | 6.431 | 26 | 229 | 54 | 14 | 2 | 2 | 4 | 5 | 108 | 1 | 0 |
| D. atra | MRAC 18560 | RWANDA | Tare, Astrida | F | 719 | 99 | 7.263 | 25 | 225 | 51 | 13 | 2 | 2 | 4 | 6 | B | 0 | |
| D. atra | MRAC 18561 | RWANDA | Tare, Astrida | F | 542 | 24 | 224 | 14 | 2 | 2 | 4 | 6 | B | 1 | 0 | |||
| D. atra | ZFMK 29567 | SOUTH SUDAN | 10 km S of Juba | F | 760 | 24 | 236 | 57 | 13 | 2 | 2 | 4 | 5 | M | ||||
| D. atra | ZFMK 29565 | SOUTH SUDAN | Aweil | F | 760 | 23 | 225 | 51 | 14 | 2 | 2 | 4 | 4 | M | ||||
| D. atra | BM 1978.379 | SOUTH SUDAN | Gilo, Imatong Mts | F | 625 | 97 | 6.443 | 24 | 229 | 13 | 2 | 4 | 4 | 6 | M | 1 | 0 | |
| D. atra | MCZ 53569 | SOUTH SUDAN | Gilo, Imatong Mts | F | 372 | 26 | 228 | 14 | 2 | 4 | 4 | 6 | B | 1 | 1 | |||
| D. atra | ZFMK 34532 | SOUTH SUDAN | Gilo, Imatong Mts | F | 24 | 228 | 63 | 2 | 2 | 4 | 6 | M | ||||||
| D. atra | ZFMK 29732 | SOUTH SUDAN | Talanga Forest | F | 500 | 24 | 227 | 54 | 14 | 2 | 2 | 4 | 6 | M | ||||
| D. atra | ZFMK 29733 | SOUTH SUDAN | Talanga Forest | F | 430 | 24 | 59 | 14 | 2 | 2 | 4 | 9 | M | |||||
| D. atra | PEM 17329 | TANZANIA | Klein’s Gate, N.E Serengeti | F | 317 | 45 | 7.044 | 27 | 240 | 56 | 14 | 2 | 4 | 92 | 1 | 0 | ||
| D. atra | PEM 17336 | TANZANIA | Klein’s Gate, N.E Serengeti | F | 434 | 25 | 227 | 14 | 2 | 4 | 100 | 1 | 0 | |||||
| D. atra | PEM 17941 | TANZANIA | Klein’s Gate, N.E Serengeti | F | 345 | 25 | 225 | 14 | 2 | 4 | 4 | 6 | 80 | 1 | 1 | |||
| D. atra | ZMUC 6140* | TANZANIA | Minziro Forest Reserve | F | 23 | 227 | 54 | 14 | 2 | 2 | 4 | 6 | M | 1 | 0 | |||
| D. atra | ZSM 81/1936c | TANZANIA | Moshi (Uru) | F | 475 | 60 | 7.917 | 23 | 235 | 47 | 14 | 2Bx | ||||||
| D. atra | ZMB 17039A* | TANZANIA | Mount Kilimanjaro | F | 197 | 30 | 6.567 | 25 | 240 | 55 | 14 | 2 | 4 | 4 | 6 | 93 | 1 | 0 |
| D. atra | ZMB 17039B | TANZANIA | Mount Kilimanjaro | F | 408 | 62 | 6.581 | 26 | 233 | 56 | 14 | 2 | 4 | 4 | 6 | 2Bx | 1 | 0 |
| D. atra | LIV 1960.115.3 | UGANDA | Bitereko, Igara | F | 638 | 96 | 6.646 | 223 | 54 | 2 | 4 | 5 | M | |||||
| D. atra | BM 51.1.3.68 | UGANDA | Bufundi, Kigezi | F | 501 | 75 | 6.68 | 24 | 222 | 55 | 14 | 2 | 2 | 108 | 1 | 0 | ||
| D. atra | BM 51.1.3.79 | UGANDA | Bufundi, Kigezi | F | 665 | 96 | 6.927 | 24 | 216 | 50 | 14 | 2 | 2 | 4 | 6 | 2Bx | 1 | 0 |
| D. atra | NMZB 217 = SRM 3.2* | UGANDA | Bufundi, Lake Bunyonyi | F | 580 | 88 | 6.591 | 25 | 221 | 52 | 14 | 2 | 2 | 4 | 6 | B | 1 | 0 |
| D. atra | BM 51.1.3.80* | UGANDA | Bukakata | F | 645 | 101 | 6.386 | 24 | 227 | 56 | 14 | 2 | 2 | 4 | 5 | B | 1 | 0 |
| D. atra | BM 1951.1.5.24* | UGANDA | Busingiro, Budango | F | 760 | 106 | 7.17 | 23 | 230 | 54 | 14 | 2 | 4 | 4 | 6 | M | 1 | 0 |
| D. atra | MCZ 40574 | UGANDA | Butandiga, Mount Elgon | F | 650 | 97 | 6.701 | 24 | 224 | 52 | 14 | 2 | 4 | 4 | 6 | 2Bx | 1 | 0 |
| D. atra | LACM 38696* | UGANDA | Bwamba Forest (Ntandi) | F | 567 | 92 | 6.163 | 25 | 226 | 55 | 14 | 2 | 2 | 4 | 6 | B | 1 | 0 |
| D. atra | LACM 46344* | UGANDA | Bwamba Forest (Ntandi) | F | 24 | 217 | 64 | 14 | 2 | 3 | 4 | 6 | B | 1 | 1 | |||
| D. atra | CAS-SU 21709 | UGANDA | Fort Portal, Kingami | F | 410 | 70 | 5.857 | 25 | 228 | 60 | 14 | 2 | 2 | 4 | 6 | 2Bx | 1 | 1 |
| D. atra | MCZ 54738* | UGANDA | Fort Portal, Kingami | F | 700 | 108 | 6.481 | 27 | 216 | 54 | 14 | 2 | 2 | 4 | 7 | M | 1 | 0 |
| D. atra | BM 51-1-3-77* | UGANDA | Gulu | F | 475 | 63 | 7.54 | 26 | 232 | 48 | 14 | 2 | 4 | 4 | 6 | 88 | 1 | 1 |
| D. atra | BM 1976.2265 | UGANDA | Ibanda | F | 390 | 63 | 6.19 | 22 | 52 | 15 | 2 | 2 | 5 | 8 | M | 1 | 2 | |
| D. atra | BM 12.10.12.2* | UGANDA | Jinja | F | 422 | 57 | 7.404 | 26 | 230 | 51 | 14 | 2 | 4 | 4 | 6 | B | 1 | 0 |
| D. atra | BM 1961.1803* | UGANDA | Kabingo, Bunyaruguru | F | 445 | 67 | 6.642 | 25 | 211 | 53 | 14 | 2 | 4 | 4 | 6 | B | 1 | 0 |
| D. atra | LIV 1960.67.5 | UGANDA | Kagando, Kigagati | F | 205 | 30 | 6.833 | 219 | 49 | 4 | 4 | 6 | 2Bx | |||||
| D. atra | LACM 60705* | UGANDA | Kalinzu Forest | F | 392 | 59 | 6.644 | 25 | 213 | 53 | 15 | 2 | 2 | 4 | 6 | M | 1 | 0 |
| D. atra | BM 1984.882* | UGANDA | Kampala | F | 710 | 119 | 5.966 | 23 | 223 | 57 | 14 | 2 | 2 | 4 | 6 | B | 1 | 0 |
| D. atra | MUZM | UGANDA | Kampala | F | 475 | 73 | 6.507 | 25 | 224 | 56 | 14 | 3 | B | |||||
| D. atra | MUZM | UGANDA | Kampala | F | 310 | 49 | 6.327 | 25 | 220 | 54 | 14 | 2 | 2 | 4 | 6 | M | ||
| D. atra | LACM 39242* | UGANDA | Katera, Sango Bay Forest | F | 668 | 100 | 6.68 | 25 | 222 | 56 | 14 | 2 | 2 | 4 | 6 | B | 1 | 0 |
| D. atra | NMZB 103 = SRM 3.1* | UGANDA | Katunguru | F | 325 | 54 | 6.019 | 25 | 216 | 56 | 14 | 2 | 2 | 4 | 8 | M | 1 | 0 |
| D. atra | MRAC 9997* | UGANDA | Katwe | F | 25 | 217 | 60 | 14 | 2 | 4 | 4 | 6 | M | 1 | 0 | |||
| D. atra | LACM 35090* | UGANDA | Kayonza, Impenetrable F. | F | 530 | 87 | 6.092 | 24 | 224 | 57 | 14 | 2 | 2 | 4 | 6 | B | 1 | 0 |
| D. atra | LIV 1962.9.27 | UGANDA | Kayonza, Kinkizi | F | 657 | 105 | 6.257 | 216 | 53 | 2 | 4 | 6 | M | |||||
| D. atra | LIV 1962.9.22 | UGANDA | Kinani, Nyabushozi | F | 222 | 2 | 4 | 4 | M | |||||||||
| D. atra | BM 51.1.3.75 | UGANDA | Kyamahunga | F | 630 | 94 | 6.702 | 24 | 225 | 55 | 14 | 2 | 2 | 4 | 6 | 2Bx | 1 | 0 |
| D. atra | LIV 1962.9.4 | UGANDA | Kyamogorani, Kashari | F | 648 | 100 | 6.48 | 226 | 55 | 2 | 4 | 6 | B | |||||
| D. atra | FMNH 9913 | UGANDA | Lake Bunyonyi | F | 527 | 76 | 6.934 | 24 | 224 | 52 | 14 | 2 | 2 | 4 | 4 | 2Bx | 1 | |
| D. atra | MCZ 34922 | UGANDA | Lake Bunyonyi | F | 628 | 100 | 6.28 | 24 | 223 | 51 | 15 | 2 | 2 | 4 | 4 | 2Bx | 1 | 0 |
| D. atra | NMZB 219 = SRM 3.4* | UGANDA | Lake Edward | F | 620 | 101 | 6.139 | 26 | 218 | 54 | 14 | 2 | 2 | 4 | 6 | B | 1 | 0 |
| D. atra | BM 08.10.20.15* | UGANDA | Mabira Forest, Chagwe | F | 391 | 59 | 6.627 | 26 | 219 | 54 | 14 | 2 | 4 | 4 | 6 | B | 1 | 0 |
| D. atra | BM 51.1.3.76* | UGANDA | Masese, Jinja | F | 754 | 121 | 6.231 | 24 | 225 | 56 | 14 | 2 | 2 | 4 | 6 | B | 1 | 0 |
| D. atra | MCZ 48359 | UGANDA | Mihunga, Ruwenzori | F | 230 | 24 | 222 | 14 | 2 | 2 | 4 | 6 | M | 1 | 0 | |||
| D. atra | NMK 3291 | UGANDA | Mityana | F | 530 | 78 | 6.795 | 232 | 14 | 4 | M | |||||||
| D. atra | BM 1954.1.12.37* | UGANDA | Morulinga, 24 km S of Moroto | F | 300 | 43 | 6.977 | 24 | 224 | 54 | 14 | 2 | 4 | 4 | 6 | 82 | 1 | 1 |
| D. atra | CAS 85300* | UGANDA | Mount Elgon | F | 620 | 90 | 6.889 | 23 | 220 | 52 | 14 | 2 | 2 | 4 | 4 | M | 1 | 0 |
| D. atra | BM 34.12.15.592* | UGANDA | Mubango, Mabira Forest | F | 830 | 133 | 6.241 | 23 | 230 | 59 | 14 | 2 | 2 | 4 | 6 | M | 1 | 0 |
| D. atra | BM 34.12.15.588 | UGANDA | Muko, Kigezi | F | 625 | 102 | 6.127 | 24 | 214 | 55 | 14 | 2 | 2 | 4 | 6 | 2Bx | 1 | 0 |
| D. atra | BM 34.12.15.594* | UGANDA | Muko, Kigezi | F | 365 | 45 | 8.111 | 24 | 222 | 52 | 14 | 2 | 2 | 4 | 6 | M | 1 | 0 |
| D. atra | BM 03-12-2-22* | UGANDA | Mulema | F | 500 | 68 | 7.353 | 25 | 226 | 51 | 14 | 2 | 4 | 4 | 6 | 92 | 1 | 0 |
| D. atra | MCZ 48362* | UGANDA | Mushongero, Lake Mutanda | F | 438 | 64 | 6.844 | 24 | 230 | 52 | 14 | 2 | 2 | 4 | 6 | B | 1 | 0 |
| D. atra | MCZ 48360 | UGANDA | Nyakabande | F | 677 | 98 | 6.908 | 25 | 226 | 53 | 14 | 2 | 2 | 5 | 10 | 2Bx | 1 | 0 |
| D. atra | MCZ 48361 | UGANDA | Nyakabande | F | 538 | 74 | 7.27 | 24 | 223 | 51 | 14 | 2 | 2 | 4 | 6 | 2Bx | 1 | 0 |
| D. atra | UMMZ 88532/2 | UGANDA | Nyakabende | F | 325 | 45 | 7.222 | 25 | 216 | 51 | 14 | 4 | 2 | 4 | 6 | 2Bx | 1 | |
| D. atra | LIV 1959.83.2 | UGANDA | Nyamuhumia, Igara | F | 365 | 49 | 7.449 | 218 | 47 | 4 | 4 | 4 | 2Bx | |||||
| D. atra | LIV 1980.150.33 | UGANDA | Nyansimbo, Mitoma | F | 222 | 38 | 5.842 | 223 | 55 | 2 | 4 | 5 | B | |||||
| D. atra | BM 1976.1849* | UGANDA | Nyarusiza, Bufumbira, Kigezi | F | 270 | 42 | 6.429 | 24 | 214 | 53 | 14 | 2 | 2 | 4 | 6 | 95 | 1 | 0 |
| D. atra | NMZB-UM 2552* | UGANDA | Nyarusiza, Bufumbira, Kigezi | F | 310 | 50 | 6.2 | 24 | 216 | 54 | 14 | 2 | 2 | 4 | 6 | 96 | 1 | 0 |
| D. atra | MCZ 54744 | UGANDA | Rucho, Ankole | F | 592 | 102 | 5.804 | 25 | 225 | 59 | 14 | 2 | 2 | 4 | 6 | 2Bx | 1 | 0 |
| D. atra | LIV 1960.67.3 | UGANDA | Ruhama, Rwampara | F | 455 | 65 | 7 | 220 | 52 | 4 | 4 | 6 | 2Bx | |||||
| D. atra | MSNG 27755A* | UGANDA | Sesse Islands, Lake Victoria | F | 616 | 76 | 8.105 | 22 | 229 | 55 | 14 | 2 | 4 | 4 | 4 | B | 1 | 0 |
| D. atra | MCZ 40564* | UGANDA | Sipi, Mount Elgon | F | 725 | 100 | 7.25 | 23 | 224 | 55 | 14 | 2 | 2 | 4 | 4 | M | 1 | 0 |
| D. atra | MCZ 40565* | UGANDA | Sipi, Mount Elgon | F | 621 | 90 | 6.9 | 24 | 218 | 52 | 14 | 2 | 2 | 4 | 4 | M | 1 | 0 |
| D. atra | MCZ 40566 | UGANDA | Sipi, Mount Elgon | F | 515 | 77 | 6.688 | 24 | 218 | 52 | 14 | 2 | 2 | 4 | 4 | 2Bx | 1 | 0 |
| D. atra | MCZ 40569* | UGANDA | Sipi, Mount Elgon | F | 710 | 106 | 6.698 | 24 | 232 | 53 | 14 | 2 | 3 | 4 | 6 | M | 1 | 0 |
| D. atra | MCZ 40571* | UGANDA | Sipi, Mount Elgon | F | 620 | 84 | 7.381 | 24 | 220 | 52 | 14 | 2 | 3 | 4 | 7 | M | 1 | 0 |
| D. atra | MCZ 40572 | UGANDA | Sipi, Mount Elgon | F | 342 | 50 | 6.84 | 218 | 50 | 14 | 2 | 4 | 4 | 4 | B | |||
| D. atra | MCZ 40573 | UGANDA | Sipi, Mount Elgon | F | 225 | 35 | 6.429 | 223 | 54 | 14 | 2 | 4 | 4 | 4 | B | |||
| D. atra | MRAC 82-16-R4 | BURUNDI | Bujumbura | U | 23 | 218 | 75 | 14 | 2 | 2 | 4 | 5 | B | 1 | 0 | |||
| D. atra | MSNM 2975 | ETHIOPIA | El Dire, Tertale | U | 277 | 42 | 6.595 | 24 | 219 | 49 | 14 | 2 | 4 | 4 | 6 | 69 | ||
| D. atra | BM 1973.3175 | ETHIOPIA | Ghimbi | U | M | |||||||||||||
| D. atra | BM 1975.2144 | ETHIOPIA | Kebre Mengist to Negele | U | 170 | 14 | 2 | 2 | 4 | 6 | 2Bx | 1 | 1 | |||||
| D. atra | NMK 1962 | KENYA | ? | U | 377 | 72 | 5.236 | 203 | 59 | 14 | 4 | 2Bx | ||||||
| D. atra | NMK 348 | KENYA | ? | U | 192 | 27 | 7.111 | 212 | 53 | 12 | 4 | B | ||||||
| D. atra | NMK 1348 | KENYA | Athi River | U | 636 | 93 | 6.839 | 221 | 50 | 14 | 4 | B | ||||||
| D. atra | NMK 488 | KENYA | Kacheliba | U | 536 | 87 | 6.161 | 229 | 54 | 14 | 2 | 98 | ||||||
| D. atra | NMK 482 | KENYA | Kakamega | U | 257 | 39 | 6.59 | 216 | 48 | 14 | 2 | B | ||||||
| D. atra | NMK 2715 | KENYA | Kakamega | U | 411 | 65 | 6.323 | 221 | 54 | 14 | 2 | 2Bx | ||||||
| D. atra | NMK 1260 | KENYA | Kericho | U | 712 | 101 | 7.05 | 216 | 51 | 16 | 4 | B | ||||||
| D. atra | NMK 481 | KENYA | Kiambu | U | 199 | 32 | 6.219 | 226 | 69 | 12 | 4 | B | ||||||
| D. atra | NMK 2952/2 | KENYA | Kiboko Ranch Station | U | 182 | 29 | 6.276 | 251 | 64 | 14 | 4 | 2Bx | ||||||
| D. atra | NMK 3123 | KENYA | Kiserian, Baringo | U | 235 | 33 | 7.121 | 227 | 51 | 14 | 4 | B | ||||||
| D. atra | NMK 3051 | KENYA | Kitale | U | 317 | 69 | 4.594 | 214 | 52 | 14 | 2 | 2Bx | ||||||
| D. atra | NMK 1791 | KENYA | Lake Baringo | U | 619 | 107 | 5.785 | 228 | 57 | 14 | 2 | 104 | ||||||
| D. atra | NMK 3152 | KENYA | Mount Elgon | U | 208 | 40 | 5.2 | 214 | 59 | 12 | 4 | 2Bx | ||||||
| D. atra | NMK 658 | KENYA | Mount Elgon | U | 565 | 90 | 6.278 | 221 | 52 | 12 | 2 | B | ||||||
| D. atra | NMK 3557 | KENYA | Nanyuki River, Laikipia | U | 539 | 91 | 5.923 | 221 | 54 | 14 | 4 | M | ||||||
| D. atra | NMK 1489 | KENYA | Nyambene Hills, Meru | U | 594 | 97 | 6.124 | 225 | 52 | 14 | 2 | 92 | 1 | 1 | ||||
| D. atra | NMK 3182 | KENYA | Timbilil, SW Mau Forest | U | 195 | 232 | 14 | 4 | B | |||||||||
| D. atra | IRSNB 2657 | RWANDA | Ruhengeri | U | 245 | 46 | 5.326 | 23 | 209 | 69 | 14 | 2 | 2 | 4 | 5 | 106 | 1 | 0 |
| D. atra | PEM 16778 | TANZANIA | Klein’s Camp, N.E. Serengeti | U | 251 | 27 | 244 | 14 | 2 | 4 | 75 | 2 | 0 | |||||
| D. atra | ZMUC 6138 | TANZANIA | Minziro Forest Reserve | U | 212 | 38 | 5.579 | 22 | 216 | 61 | 14 | 2 | 2 | 4 | 6 | M | 1 | 0 |
| D. atra | ZMUC 6141 | TANZANIA | Minziro Forest Reserve | U | 23 | 214 | 68 | 12 | 2 | 2 | 4 | 6 | B | 1 | 0 | |||
| D. atra | BM 1970.2178 | TANZANIA | Sitete, Mahale Peninsula | U | 435 | 43 | 64 | 14 | 2 | 2 | 4 | 4 | B | 1 | 0 | |||
| D. atra | MSNG 30413A | UGANDA | Bussu, Jinja | U | 786 | 102 | 7.706 | 25 | 230 | 60 | 14 | 2 | 4 | 4 | 6 | M | 1 | 0 |
| D. atra | MSNG 52410 | UGANDA | Bussu, Jinja | U | 586 | 71 | 8.254 | 23 | 226 | 54 | 12 | 2 | 4 | 4 | B | |||
| D. atra | LIV 1960.115.4 | UGANDA | Kayonza, Kinkizi | U | 2 | 4 | 5 | B | ||||||||||
| D. atra | NMK 1495 | UGANDA | Nyarusiza | U | 664 | 93 | 7.14 | 226 | 50 | 14 | 2 | |||||||
| D. atra | NMK 1496 | UGANDA | Nyarusiza | U | 591 | 94 | 6.287 | 221 | 59 | 14 | 2 | B | ||||||
| D. bazi | NMW 26879/1 | EGYPT | Cairo, surroundings of | F | 461 | 61 | 7.557 | 24 | 234 | 50 | 14 | 2 | 4 | 4 | 6 | 1 | 1 | |
| D. bazi | ZMB 1861* | EGYPT | Oberaegypten | M | 455 | 79 | 5.759 | 23 | 229 | 59 | 14 | 2 | 4 | 4 | 6 | 48 | 1 | 1 |
| D. bazi | MNHN 1847* | EGYPT | M | 410 | 70 | 5.857 | 23 | 229 | 59 | 14 | 2 | 4 | 4 | 6 | 49 | 2 | 1 | |
| D. bazi | ZMB 5865* | EGYPT | M | 427 | 78 | 5.474 | 23 | 219 | 60 | 14 | 2 | 4 | 6 | 6 | 39 | 2 | 2 | |
| D. bazi | BM 97.10.28.575* | EGYPT | Fayoum | F | 602 | 90 | 6.689 | 24 | 227 | 55 | 14 | 2 | 4 | 5 | 6 | 42 | 1 | 1 |
| D. bazi | ZMB 51552* | EGYPT | Uberegypte | F | 503 | 70 | 7.186 | 24 | 228 | 51 | 14 | 2 | 4 | 4 | 6 | 48 | 0 | 0 |
| D. cf. bazi | ZFMK 38415 | SUDAN | Erkowit | M | 25 | 229 | 67 | 14 | 2 | 4 | 6 | 8 | 48 | 1 | ||||
| D. confusa | NMW 9964/2* | ETHIOPIA | ? | M | 362 | 67 | 5.403 | 24 | 208 | 57 | 14 | 2 | 4 | 4 | 7 | 75 | 1 | 0 |
| D. confusa | ZMB 56436* | ETHIOPIA | ‘Addis Ababa’ | M | 475 | 87 | 5.46 | 24 | 213 | 57 | 14 | 2 | 4 | 4 | 6 | 79 | 0 | 1 |
| D. confusa | NMK 485 | KENYA | Kacheliba | M | 319 | 55 | 5.8 | 212 | 51 | 14 | 4 | 67 | ||||||
| D. confusa | NMK 489 | KENYA | Kacheliba | M | 424 | 74 | 5.73 | 216 | 54 | 14 | 4 | 65 | ||||||
| D. confusa | MCZ 53403* | SOUTH SUDAN | Nimule | M | 232 | 42 | 5.524 | 26 | 208 | 63 | 14 | 2 | 4 | 4 | 6 | 55 | 1 | 1 |
| D. confusa | FMNH 58361 | SOUTH SUDAN | Torit | M | 396 | 79 | 5.013 | 25 | 224 | 67 | 14 | 2 | 4 | 4 | 6 | B | 1 | |
| D. confusa | FMNH 58362 | SOUTH SUDAN | Torit | M | 330 | 69 | 4.783 | 23 | 210 | 63 | B | 1 | ||||||
| D. confusa | FMNH 62209 | SOUTH SUDAN | Torit | M | 551 | 25 | 203 | 14 | 2 | 4 | 4 | 6 | B | 1 | ||||
| D. confusa | FMNH 62214 | SOUTH SUDAN | Torit | M | 394 | 78 | 5.051 | 24 | 220 | 64 | 14 | 2 | 4 | 4 | 5 | 54 | 1 | |
| D. confusa | FMNH 62217 | SOUTH SUDAN | Torit | M | 235 | 45 | 5.222 | 25 | 212 | 64 | 14 | 2 | 4 | 4 | 6 | B | 1 | |
| D. confusa | MUZM 10/R1* | UGANDA | Ajai Game Reserve | M | 395 | 84 | 4.702 | 25 | 200 | 66 | 14 | 2 | 4 | 4 | 8 | 50 | 1 | 0 |
| D. confusa | MCZ 47815* | UGANDA | Busingiro, Budongo Forest | M | 358 | 64 | 5.594 | 26 | 199 | 54 | 14 | 2 | 4 | 4 | 10 | 54 | 1 | 1 |
| D. confusa | BM 51-1-5-39* | UGANDA | Lira, Lango | M | 325 | 62 | 5.242 | 25 | 205 | 62 | 14 | 2 | 4 | 4 | 6 | 62 | 1 | 0 |
| D. confusa | UMMZ 88529 | UGANDA | Lira, Lango | M | 45 | 25 | 14 | 2 | 4 | 4 | 7 | 61 | 0 | |||||
| D. confusa | LIV 1960.51.2 | UGANDA | Nakosa, Bunyaruguru | M | 390 | 208 | 4 | 4 | 6 | |||||||||
| D. confusa | CAS 141784* | KENYA | Mumias | F | 203 | 27 | 7.519 | 26 | 226 | 48 | 14 | 2 | 4 | 4 | 6 | 63 | 1 | 0 |
| D. confusa | ZMB 77441 | RWANDA | Lac Kivu | F | 620 | 24 | 224 | 11 | 2 | 4 | 4 | 6 | 71 | 1 | 0 | |||
| D. confusa | ZFMK 29694 | SOUTH SUDAN | 10km S of Juba | F | 25 | 55 | 14 | 2 | 4 | 4 | 6 | 50 | ||||||
| D. confusa | FMNH 58496 | SOUTH SUDAN | Juba | F | 575 | 92 | 6.25 | 26 | 228 | 54 | 14 | 2 | 4 | 4 | 8 | 54 | 1 | |
| D. confusa | MCZ 53401* | SOUTH SUDAN | Logutoic | F | 77 | 26 | 221 | 55 | 14 | 2 | 4 | 4 | 6 | 60 | 1 | 0 | ||
| D. confusa | FMNH 62216 | SOUTH SUDAN | Lokwi | F | 165 | 25 | 6.6 | 25 | 229 | 49 | 14 | 2 | 3 | 4 | 6 | 69 | 1 | |
| D. confusa | MCZ 53402 | SOUTH SUDAN | Mongalla | F | 524 | 23 | 225 | 14 | 2 | 4 | 4 | 6 | 58 | 1 | 0 | |||
| D. confusa | NMW 9964/7+ | SOUTH SUDAN | ‘Sudan’ (taken as East Equatoria) | F | 465 | 67 | 6.940 | 25 | 231 | 55 | 14 | 2 | 4 | 4 | 4 | 62 | 1 | 0 |
| D. confusa | FMNH 58363 | SOUTH SUDAN | Torit | F | 492 | 72 | 6.833 | 25 | 223 | 54 | 14 | 2 | 4 | 4 | 6 | |||
| D. confusa | FMNH 62207 | SOUTH SUDAN | Torit | F | 369 | 25 | 239 | 14 | 2 | 4 | 4 | 6 | 61 | 1 | ||||
| D. confusa | FMNH 62208 | SOUTH SUDAN | Torit | F | 428 | 25 | 236 | 14 | 2 | 4 | 4 | 6 | B | 1 | ||||
| D. confusa | FMNH 62210 | SOUTH SUDAN | Torit | F | 417 | 64 | 6.516 | 24 | 230 | 55 | 14 | 2 | 4 | 4 | 6 | 1 | ||
| D. confusa | FMNH 62211 | SOUTH SUDAN | Torit | F | 187 | 32 | 5.844 | 25 | 233 | 56 | 14 | 2 | 4 | 4 | 6 | B | 1 | |
| D. confusa | FMNH 62212 | SOUTH SUDAN | Torit | F | 435 | 62 | 7.016 | 25 | 238 | 53 | 14 | 2 | 4 | 4 | 6 | 68 | 1 | |
| D. confusa | FMNH 62213 | SOUTH SUDAN | Torit | F | 378 | 25 | 237 | 14 | 2 | 4 | 4 | 5 | 64 | 1 | ||||
| D. confusa | MCZ 53405* | SOUTH SUDAN | Torit | F | 324 | 51 | 6.353 | 25 | 236 | 57 | 14 | 2 | 4 | 4 | 6 | 73 | 1 | 1 |
| D. confusa | MCZ 53404 | SOUTH SUDAN | Torit | F | 546 | 24 | 236 | 14 | 2 | 4 | 4 | 6 | 64 | 1 | 1 | |||
| D. confusa | FMNH 58336 | SOUTH SUDAN | Yei | F | 252 | 51 | 4.941 | 27 | 226 | 58 | 13 | 2 | 4 | 4 | 7 | 58 | ||
| D. confusa | BM 51.1.3.69* | UGANDA | Bisu, Kigezi | F | 518 | 79 | 6.557 | 26 | 222 | 56 | 14 | 2 | 4 | 4 | 6 | 61 | 1 | 0 |
| D. confusa | LACM 39043* | UGANDA | Bugoma Forest | F | 166 | 31.5 | 5.188 | 23 | 213 | 61 | 14 | 2 | 4 | 4 | 6 | 61 | 1 | 0 |
| D. confusa | MCZ 47830* | UGANDA | Gulu, Acholi | F | 301 | 52 | 5.788 | 23 | 212 | 56 | 14 | 2 | 4 | 4 | 6 | B | 1 | 0 |
| D. confusa | NMZB 14333* | UGANDA | Kyambura Game Reserve | F | 520 | 73 | 7.123 | 25 | 229 | 52 | 14 | 2 | 4 | 4 | 8 | 63 | 1 | 0 |
| D. confusa | NMZB 15411* | UGANDA | Kyambura Game Reserve | F | 485 | 73 | 6.644 | 26 | 215 | 52 | 14 | 2 | 4 | 4 | 5 | B | 1 | 0 |
| D. confusa | MCZ 47801* | UGANDA | Lira, Lango | F | 444 | 59 | 7.525 | 25 | 228 | 50 | 14 | 2 | 4 | 4 | 8 | B | 1 | 0 |
| D. confusa | UMMZ 88530 | UGANDA | Lira, Lango | F | 56 | 27 | 218 | 50 | 14 | 2 | 4 | 4 | 8 | B | 1 | |||
| D. confusa | BM 51.1.3.84* | UGANDA | Moyo, West Madi | F | 245 | 37 | 6.622 | 26 | 229 | 57 | 14 | 2 | 4 | 4 | 6 | 55 | 1 | 0 |
| D. confusa | LIV 1962.106.19 | UGANDA | Nyanshimbo, Ibanda | F | 501 | 226 | 4 | 4 | 7 | |||||||||
| D. confusa | LIV 1960.67.2 | UGANDA | Sanga, Nyabushozi | F | 481 | 73 | 6.589 | 213 | 53 | 4 | 4 | 7 | ||||||
| D. confusa | BM 1954 1.12.8* | UGANDA | Semliki | F | 503 | 72 | 6.986 | 24 | 218 | 52 | 12 | 2 | 4 | 3 | 6 | 59 | 1 | 0 |
| D. confusa | BM 1954 1.12.9* | UGANDA | Semliki | F | 77 | 26 | 215 | 55 | 14 | 2 | 4 | 3 | 6 | 55 | 1 | 2 | ||
| D. confusa | BM 1976.848* | UGANDA | Serere, Teso | F | 567 | 81 | 7 | 26 | 227 | 55 | 14 | 2 | 4 | 4 | 5 | B | 1 | 0 |
| D. crucifera | MZUT 3582* | ERITREA | Agordat | M | 211 | 39 | 5.41 | 22 | 226 | 61 | 14 | 2 | 4 | 4 | 5 | 52 | 1 | 0 |
| D. crucifera | ZMB 7631* | ERITREA | Bogos | F | 442 | 64 | 6.906 | 25 | 231 | 52 | 14 | 2 | 4 | 4 | 6 | 62 | 1 | 0 |
| D. crucifera | MZUT 3469 | ERITREA | Agordat | F | 23 | 247 | 50 | |||||||||||
| D. crucifera | MSNG 6738* | ERITREA | Keren | F | 368 | 45 | 8.178 | 24 | 243 | 49 | 14 | 2 | 4 | 4 | 6 | 51 | 1 | 0 |
| D. fasciata | LACM 38697 | UGANDA | Ntandi, Bwamba Forest | M | 525 | 111 | 4.73 | 23 | 230 | 71 | 14 | 2 | 4 | 4 | 7 | 2 | 0 | |
| D. fasciata | LACM 46342* | UGANDA | Ntandi, Bwamba Forest | M | 509 | 116 | 4.388 | 23 | 229 | 76 | 14 | 2 | 4 | 4 | 6 | 141 | 1 | 0 |
| D. fasciata | LACM 39243* | UGANDA | Sango Bay Forest, Katera | M | 472 | 91 | 5.187 | 24 | 212 | 59 | 14 | 2 | 4 | 4 | 4 | 108 | 1 | 0 |
| D. fasciata | MCZ 48357 | UGANDA | Bundibugyo, Bwamba | F | 667 | 119 | 5.605 | 24 | 241 | 68 | 13 | 4 | 4 | 4 | 6 | 1 | 0 | |
| D. fasciata | MCZ 48358A | UGANDA | Bundibugyo, Bwamba | F | 730 | 132 | 5.53 | 23 | 239 | 67 | 14 | 2 | 4 | 4 | 6 | 1 | 0 | |
| D. fasciata | MCZ 48358B | UGANDA | Bundibugyo, Bwamba | F | 635 | 100 | 6.35 | 25 | 248 | 66 | 14 | 2 | 3 | 4 | 6 | 0 | ||
| D. fasciata | UMMZ 88531 | UGANDA | Bundibugyo, Bwamba | F | 627 | 107 | 5.86 | 23 | 248 | 64 | 14 | 4 | 4 | 4 | 6 | 1 | ||
| D. fasciata | LACM 46341 | UGANDA | Ntandi, Bwamba Forest | F | 238 | 50 | 4.76 | 22 | 237 | 76 | 14 | 2 | 2 | 6 | 7 | 1 | 0 | |
| D. fasciata | LACM 46343* | UGANDA | Ntandi, Bwamba Forest | F | 586 | 113 | 5.186 | 24 | 241 | 71 | 14 | 2 | 4 | 5 | 8 | 116 | 1 | 0 |
| D. fasciata | LACM 46345* | UGANDA | Ntandi, Bwamba Forest | F | 312 | 57 | 5.474 | 24 | 242 | 71 | 14 | 2 | 4 | 4 | 6 | 111 | 1 | 0 |
| D. fasciata | LACM 46346 | UGANDA | Ntandi, Bwamba Forest | F | 598 | 103 | 5.806 | 24 | 245 | 65 | 13 | 2 | 4 | 5 | 6 | 1 | 0 | |
| D. fasciata | TCWC 23839* | UGANDA | Ntandi, Bwamba Forest | F | 611 | 108 | 5.657 | 23 | 243 | 67 | 14 | 2 | 4 | 4 | 7 | 113 | 1 | 0 |
| D. medici | NMK 2631 | KENYA | Gede | M | 679 | 146 | 4.651 | 220 | 78 | 14 | 4 | L | ||||||
| D. medici | NMK 3200 | KENYA | Gede | M | 384 | 91 | 4.22 | 235 | 80 | 14 | 2 | L | ||||||
| D. medici | BM 51.1.3.73* | KENYA | Kilifi | M | 465 | 116 | 4.009 | 24 | 229 | 87 | 14 | 2 | 4 | 4 | 6 | L | 2 | 0 |
| D. medici | MCZ 40581* | KENYA | Lamu Island | M | 502 | 132 | 3.803 | 23 | 226 | 94 | 14 | 2 | 4 | 4 | 6 | L | 2 | 0 |
| D. medici | NMK 1561 | KENYA | Malindi | M | 487 | 127 | 3.835 | 230 | 84 | 14 | 4 | L | ||||||
| D. medici | NMK 2535 | KENYA | Malindi | M | 560 | 125 | 4.48 | 220 | 75 | 12 | 4 | L | ||||||
| D. medici | NMK 2558 | KENYA | Malindi | M | 241 | 79 | 14 | 2 | L | |||||||||
| D. medici | NMK 3292 | KENYA | Malindi | M | 616 | 133 | 4.632 | 241 | 80 | 14 | 4 | L | ||||||
| D. medici | ZMB 50191 | KENYA | Malindi | M | 461 | 115 | 4.009 | 24 | 239 | 91 | 14 | 2 | 4 | 4 | 5 | P | 1 | 0 |
| D. medici | NMK 3956 | KENYA | Mount Warges, Wamba | M | 466 | 106 | 4.396 | 24 | 239 | 83 | 14 | 2 | 4 | 4 | 6 | P | 2 | 0 |
| D. medici | NMK 1699 | KENYA | Nyambene Hills, Meru | M | 424 | 104 | 4.077 | 233 | 14 | 2 | 4 | 4 | 6 | P | 2 | 0 | ||
| D. medici | CAS 135751 | KENYA | Tiwi, Kwale District | M | 370 | 85 | 4.353 | 24 | 224 | 84 | 14 | 2 | 4 | 4 | 6 | P | 2 | 1 |
| D. medici | MZUF 2028 | SOMALIA | Alessandra Island, Gelib | M | 422 | 107 | 3.944 | 24 | 233 | 93 | 12 | 2 | 4 | 4 | 6 | 1 | 1 | |
| D. medici | CAS 148077* | SOMALIA | Mareri, Lower Juba River | M | 375 | 95 | 3.947 | 24 | 230 | 89 | 14 | 2 | 4 | 4 | 6 | L | 2 | 1 |
| D. medici | MZUF 27155 | SOMALIA | Mareri, Lower Juba River | M | 496 | 137 | 3.62 | 24 | 231 | 98 | 13 | 2 | 4 | 4 | 6 | 2 | ||
| D. medici | NMW 9964/18+ | TANZANIA | Dar-es-Salaam | M | 430 | 104 | 4.135 | 24 | 246 | 94 | 14 | 3 | 4 | 4 | 6 | 66 | 2 | 0 |
| D. medici | FMNH 251325 | TANZANIA | East Usambara Mts | M | 400 | 95 | 4.211 | 24 | 249 | 89 | 14 | 2 | 3 | 4 | 5 | P | 2 | 0 |
| D. medici | ZMUC 6144* | TANZANIA | Longido | M | 522 | 129 | 4.047 | 24 | 239 | 89 | 14 | 2 | 4 | 4 | 6 | 79 | 2 | 1 |
| D. medici | SMF 19460* | TANZANIA | Mafia Island | M | 471 | 119 | 3.958 | 23 | 252 | 98 | 14 | 2 | 4 | 4 | 7 | 68 | 2 | 0 |
| D. medici | MCZ 18206* | TANZANIA | Makindu River | M | 525 | 130 | 4.038 | 24 | 240 | 89 | 14 | 2 | 4 | 4 | 6 | 67 | 2 | 0 |
| D. medici | NMZB 17318 | TANZANIA | Mlinga Forest Res. | M | 460 | 124 | 3.71 | 23 | 240 | 103 | 14 | 2 | 4 | 4 | 6 | P | 2 | 0 |
| D. medici | USNM 62904 | TANZANIA | Morogoro | M | 475 | 115 | 4.13 | 24 | 237 | 83 | 14 | 2 | 4 | 5 | 7 | 67 | 2 | |
| D. medici | USNM 62906 | TANZANIA | Morogoro | M | 470 | 105 | 4.476 | 24 | 248 | 86 | 14 | 2 | 4 | 4 | 6 | P | 2 | |
| D. medici | SMNS 4216E | TANZANIA | Moshi | M | 26 | 230 | 14 | 2 | 4 | 4 | 6 | 72 | 2 | 0 | ||||
| D. medici | SMNS 4216F | TANZANIA | Moshi | M | 315 | 69 | 4.565 | 26 | 232 | 80 | 15 | 2 | 4 | 4 | 7 | 92 | 1 | |
| D. medici | ZMUC 6121* | TANZANIA | Msolwa, Ruhebo Mtns | M | 444 | 117 | 3.795 | 25 | 234 | 81 | 14 | 2 | 2 | 4 | 6 | 70 | 2 | 0 |
| D. medici | ZMB 17453* | TANZANIA | Usambara | M | 575 | 158 | 3.639 | 25 | 242 | 109 | 14 | 2 | 4 | 6 | 10 | 64 | 2 | 0 |
| D. medici | ZMB 8597* | TANZANIA | Zanzibar Coast | M | 480 | 117 | 4.103 | 24 | 243 | 94 | 14 | 3 | 4 | 5 | 6 | 69 | 2 | 0 |
| D. medici | NMK 3249 | TANZANIA | Zanzibar Island | M | 451 | 129 | 3.496 | 235 | 93 | 12 | 2 | |||||||
| D. medici | NMK 402 | KENYA | Chyulu Hills | F | 616 | 120 | 5.133 | 239 | 66 | 14 | 4 | |||||||
| D. medici | AMNH 88631 | KENYA | Gede | F | 641 | 130 | 4.931 | 25 | 235 | 80 | 14 | 2 | 3 | 4 | 6 | 2 | 0 | |
| D. medici | NMK 2597 | KENYA | Gede | F | 600 | 141 | 4.255 | 229 | 82 | 14 | 2 | L | ||||||
| D. medici | NMK 2619* | KENYA | Gede | F | 580 | 123 | 4.715 | 26 | 227 | 81 | 14 | 2 | 4 | 6 | 7 | L | 2 | 0 |
| D. medici | NMK 480 | KENYA | Giriama, Kilifi | F | 466 | 103 | 4.524 | 226 | 72 | 14 | 4 | |||||||
| D. medici | ZMB 37600 | KENYA | Kibognoto – Kilimanjaro | F | 496 | 103 | 4.816 | 27 | 218 | 74 | 14 | 2 | 4 | 6 | 8 | 1 | 0 | |
| D. medici | NMK 1050 | KENYA | Kikambala, Kilifi | F | 612 | 112 | 5.464 | 231 | 79 | 14 | 4 | |||||||
| D. medici | BM 51.1.3.72* | KENYA | Kilifi | F | 600 | 119 | 5.042 | 25 | 228 | 80 | 14 | 2 | 3 | 4 | 6 | L | 2 | 0 |
| D. medici | MCZ 40582* | KENYA | Lamu Island | F | 575 | 130 | 4.423 | 24 | 226 | 84 | 14 | 2 | 4 | 4 | 6 | L | 2 | 0 |
| D. medici | NMK 2522 | KENYA | Malindi | F | 567 | 141 | 4.021 | 229 | 80 | 14 | 2 | L | ||||||
| D. medici | NMK 2526 | KENYA | Malindi | F | 707 | 131 | 5.397 | 235 | 77 | 14 | 4 | L | ||||||
| D. medici | NMK 2527 | KENYA | Malindi | F | 545 | 124 | 4.395 | 247 | 85 | 14 | 4 | L | ||||||
| D. medici | NMK 2529 | KENYA | Malindi | F | 654 | 127 | 5.15 | 232 | 76 | 14 | 4 | L | ||||||
| D. medici | NMK 2530 | KENYA | Malindi | F | 616 | 148 | 4.162 | 234 | 79 | 14 | 4 | L | ||||||
| D. medici | NMK 2531 | KENYA | Malindi | F | 597 | 134 | 4.455 | 225 | 82 | 14 | 4 | L | ||||||
| D. medici | NMK 2532 | KENYA | Malindi | F | 494 | 116 | 4.259 | 227 | 79 | 14 | 4 | L | ||||||
| D. medici | NMK 2533 | KENYA | Malindi | F | 535 | 117 | 4.573 | 220 | 76 | 14 | 2 | L | ||||||
| D. medici | NMK 2587 | KENYA | Malindi | F | 566 | 128 | 4.422 | 239 | 75 | 14 | 4 | L | ||||||
| D. medici | NMK 3290 | KENYA | Malindi | F | 597 | 121 | 4.934 | 234 | 79 | 14 | 2 | L | ||||||
| D. medici | BM 98.1.8.15* | KENYA | Maungu | F | 452 | 92 | 4.913 | 25 | 228 | 81 | 14 | 2 | 4 | 5 | 8 | L | 2 | 0 |
| D. medici | NMW 26926+ | KENYA | Mombasa, 30 km S of | F | 289 | 66 | 4.379 | 26 | 233 | 81 | 14 | 2 | 4 | 4 | 6 | 86 | 2 | 0 |
| D. medici | BM 87.11.3.31* | KENYA | Mount Kilimanjaro | F | 500 | 104 | 4.808 | 24 | 230 | 80 | 14 | 2 | 4 | 5 | 6 | L | 2 | 0 |
| D. medici | USNM 16755* | KENYA | Mount Kilimanjaro | F | 702 | 25 | 232 | 67 | 13 | 2 | 4 | 7 | 6 | L | 2 | 0 | ||
| D. medici | MCZ 40580 | KENYA | Mount Mbololo | F | 645 | 120 | 5.375 | 26 | 231 | 72 | 14 | 2 | 4 | 6 | 9 | L | ||
| D. medici | ZFMK 63582 | KENYA | near Mombasa | F | 613 | 119 | 5.151 | 25 | 231 | 74 | 14 | 2 | 4 | 4 | 6 | 2 | 0 | |
| D. medici | NMK 2377 | KENYA | Nguni, Mwingi | F | 481 | 84 | 5.726 | 223 | 82 | 14 | 4 | |||||||
| D. medici | ZMB 9244* | KENYA | Taita | F | 105 | 25 | 232 | 73 | 14 | 2 | 3 | 4 | 7 | L | 2 | 0 | ||
| D. medici | ZMB 17496 | KENYA | Takaungu | F | 405 | 77 | 5.26 | 25 | 238 | 74 | 14 | 2 | 4 | 6 | 8 | 2 | 1 | |
| D. medici | FMNH 142635* | KENYA | ‘Vicinity of Nairobi’ | F | 912 | 149 | 6.121 | 25 | 235 | 71 | 14 | 2 | 4 | 6 | 8 | L | 2 | 1 |
| D. medici | BM 98.1.8.14 | KENYA | Voi to Ndi | F | 274 | 52 | 5.269 | 24 | 241 | 75 | 14 | 2 | 4 | 4 | 6 | P | 2 | 0 |
| D. medici | MSNG 42710b | SOMALIA | Belet Amin | F | 845 | 22 | 232 | 13 | 2 | 4 | 4 | 6 | L | 0 | 0 | |||
| D. medici | MCZ 23274* | TANZANIA | Amani, Usambara Mts | F | 640 | 145 | 4.414 | 26 | 241 | 87 | 14 | 2 | 4 | 4 | 6 | 108 | 2 | 0 |
| D. medici | ZMB 37598* | TANZANIA | Amani, Usambara Mts | F | 546 | 117 | 4.667 | 26 | 241 | 82 | 14 | 2 | 4 | 4 | 7 | 77 | 2 | 0 |
| D. medici | ZMUC 6123 | TANZANIA | Bombani, E. Usambara Mts | F | 649 | 124 | 5.234 | 25 | 239 | 73 | 14 | 2 | 4 | 4 | 6 | P | 2 | 0 |
| D. medici | ZMUC 6133* | TANZANIA | Bombani, E. Usambara Mts | F | 599 | 124 | 4.831 | 25 | 245 | 78 | 14 | 2 | 4 | 4 | 6 | 76 | 2 | 0 |
| D. medici | NMZB 11265* | TANZANIA | Mafia Island | F | 410 | 87 | 4.713 | 26 | 252 | 88 | 14 | 2 | 4 | 4 | 6 | L | 2 | 0 |
| D. medici | AMNH 143717 | TANZANIA | Morogoro | F | 262 | 54 | 4.852 | 24 | 243 | 83 | 14 | 2 | 4 | 4 | 6 | 64 | 2 | 2 |
| D. medici | USNM 62901 | TANZANIA | Morogoro | F | 595 | 118 | 5.042 | 25 | 240 | 81 | 14 | 2 | 4 | 4 | 6 | 59 | 2 | |
| D. medici | ZMB 27616* | TANZANIA | Morogoro | F | 630 | 120 | 5.25 | 26 | 237 | 72 | 14 | 2 | 4 | 4 | 6 | 70 | 2 | 0 |
| D. medici | SMNS 4216A* | TANZANIA | Moshi | F | 680 | 135 | 5.037 | 24 | 237 | 75 | 15 | 2 | 4 | 5 | 7 | L | 2 | 1 |
| D. medici | SMNS 4216B* | TANZANIA | Moshi | F | 117 | 25 | 233 | 69 | 14 | 2 | 4 | 5 | 8 | L | 2 | 0 | ||
| D. medici | SMNS 4216C* | TANZANIA | Moshi | F | 500 | 93 | 5.376 | 26 | 233 | 61 | 14 | 2 | 4 | 5 | 7 | L | 1 | 0 |
| D. medici | SMNS 4216D* | TANZANIA | Moshi | F | 419 | 82 | 5.11 | 25 | 232 | 75 | 14 | 2 | 4 | 6 | 8 | L | 2 | 0 |
| D. medici | ZFMK 82342 | TANZANIA | Mount Kwaraha | F | 287 | 25 | 235 | 14 | 2 | 4 | 4 | 6 | 78 | 1 | 0 | |||
| D. medici | MCZ 23273* | TANZANIA | Nyange, Uluguru Mts | F | 463 | 94 | 4.926 | 25 | 244 | 82 | 14 | 2 | 4 | 4 | 6 | 57 | 2 | 0 |
| D. medici | CM 36586 | TANZANIA | Usa River, Arusha | F | 460 | 85 | 5.412 | 26 | 232 | 75 | 14 | 2 | 4 | 4 | 6 | L | ||
| D. medici | BM 50.1.5.40 | TANZANIA | Zanzibar | F | 650 | 131 | 4.962 | 25 | 247 | 77 | 14 | 2 | 4 | 4 | 6 | P | 2 | 0 |
| D. medici | ZMB 17219* | TANZANIA | Zanzibar | F | 568 | 112 | 5.071 | 25 | 242 | 77 | 14 | 2 | 4 | 6 | 8 | 82 | 2 | 0 |
| D. medici | ZMB 5737* | TANZANIA | Zanzibar | F | 595 | 133 | 4.474 | 25 | 247 | 90 | 14 | 2 | 4 | 5 | 8 | 80 | 2 | 0 |
| D. medici | NMK 4121 | KENYA | Kitobo Forest, Taveta | U | 672 | 106 | 6.34 | 233 | 66 | 14 | 4 | |||||||
| D. medici | NMK 2538 | KENYA | Malindi | U | 597 | 134 | 4.455 | 231 | 70 | 2 | L | |||||||
| D. medici | NMK 2540 | KENYA | Malindi | U | 422 | 123 | 3.431 | 229 | 79 | 4 | P | |||||||
| D. medici | NMK 4131 | KENYA | River Tana, Mchelelo Forest | U | 484 | 83 | 5.831 | 232 | 84 | 12 | 4 | |||||||
| D. medici | ZMUC 6124 | TANZANIA | Amani, E. Usambara Mts | U | 330 | 73 | 4.521 | 26 | 246 | 84 | 14 | 2 | 4 | 4 | 8 | 91 | 2 | |
| D. medici | ZMUC 6143 | TANZANIA | East Usambara Mts | U | 182 | 46 | 3.957 | 230 | 92 | 14 | 4 | 4 | 4 | 5 | 59 | 1 | 1 | |
| D. medici | ZMB 48150 | TANZANIA | Kimboza Forest | U | 241 | 14 | 2 | 4 | 4 | 4 | 75 | |||||||
| D. medici | MCZ 54827 | TANZANIA | Mafia Island | U | 14 | 2 | 4 | 6 | 7 | P | 2 | 1 | ||||||
| D. medici | ZMUC 6111 | TANZANIA | Zanzibar Island | U | 382 | 78 | 4.897 | 25 | 251 | 83 | 14 | 2 | 4 | 4 | 6 | P | 2 | 0 |
| D. scabra | PEM 8602* | ETHIOPIA | Entrance to Awash Nat. Park | M | 385 | 71 | 5.423 | 23 | 212 | 57 | 14 | 2 | 4 | 5 | 6 | 51 | 2 | 2 |
| D. scabra | BM 1973.3259* | ETHIOPIA | Lake Abiata | M | 435 | 80 | 5.438 | 24 | 215 | 56 | 14 | 2 | 4 | 4 | 6 | 75 | 1 | 1 |
| D. scabra | ZMB 51835* | ETHIOPIA | Meki | M | 420 | 70 | 6 | 23 | 214 | 57 | 16 | 2 | 4 | 4 | 7 | 77 | 1 | 0 |
| D. scabra | BM 00.11.29.3 | ETHIOPIA | west of Omo River | M | 445 | 24 | 212 | 14 | 2 | 4 | 4 | 6 | 60 | 2 | 0 | |||
| D. scabra | AAU 489 | ETHIOPIA | west shore of Lake Chamo | M | 313 | 52 | 6.019 | 25 | 218 | 62 | 14 | 2 | 4 | 4 | 4 | |||
| D. scabra | MSNPV CR790* | KENYA | Gatab, Mount Kulal | M | 384 | 81 | 4.741 | 24 | 220 | 63 | 14 | 2 | 4 | 4 | 6 | 92 | 1 | 1 |
| D. scabra | MSNPV CR792* | KENYA | Gatab, Mount Kulal | M | 312 | 64 | 4.875 | 24 | 219 | 66 | 14 | 2 | 4 | 4 | 6 | 84 | 1 | 2 |
| D. scabra | BM 01.1.3.3* | KENYA | Gilgil | M | 398 | 79 | 5.038 | 26 | 208 | 65 | 14 | 2 | 2 | 4 | 7 | 76 | 2 | 0 |
| D. scabra | CAS 130785* | KENYA | Isiolo | M | 385 | 75 | 5.133 | 25 | 203 | 60 | 14 | 2 | 4 | 4 | 6 | 79 | 1 | 0 |
| D. scabra | NMK 561 | KENYA | Isiolo | M | 386 | 71 | 5.437 | 220 | 59 | 14 | 4 | 81 | ||||||
| D. scabra | BM 93.11.21.72* | KENYA | Kithu-Uri, E. Kikuyu | M | 350 | 59 | 5.932 | 25 | 206 | 54 | 14 | 2 | 4 | 4 | 6 | 66 | 1 | 0 |
| D. scabra | NMK 2965 | KENYA | Kora | M | 456 | 88 | 5.182 | 221 | 62 | 14 | 4 | 65 | ||||||
| D. scabra | NMK 1472 | KENYA | Lake Naivasha | M | 367 | 63 | 5.825 | 213 | 61 | 14 | 4 | 80 | ||||||
| D. scabra | NMK 2343 | KENYA | Lake Naivasha | M | 412 | 84 | 4.905 | 206 | 57 | 14 | 4 | 69 | ||||||
| D. scabra | NMK 3052 | KENYA | Solio Ranch, Laikipia | M | 288 | 54 | 5.333 | 204 | 50 | 14 | 4 | 66 | ||||||
| D. scabra | NMK 3021 | KENYA | Tsavo East N.P. | M | 236 | 41 | 5.756 | 208 | 47 | 14 | 4 | 57 | ||||||
| D. scabra | NMK 1022 | KENYA | Voi | M | 298 | 52 | 5.731 | 208 | 52 | 14 | 4 | 73 | ||||||
| D. scabra | PEM 5624 | RWANDA | 15 km S of Butare | M | 351 | 76 | 4.618 | 25 | 197 | 67 | 14 | 2 | 4 | 4 | 7 | 1 | 1 | |
| D. scabra | MRAC 18559 | RWANDA | Astrida | M | 136 | 25 | 192 | 14 | 2 | 4 | 4 | 5 | 61 | 1 | ||||
| D. scabra | MRAC 9209* | RWANDA | Gabiro | M | 192 | 35 | 5.486 | 26 | 201 | 63 | 14 | 2 | 4 | 4 | 7 | 58 | 1 | 2 |
| D. scabra | ZMB 22518* | RWANDA | Lake Kivu | M | 420 | 108 | 3.889 | 24 | 201 | 73 | 12 | 2 | 3 | 4 | 7 | 64 | 1 | 0 |
| D. scabra | MZUF 24179 | SOMALIA | 6 km E of Afgoi | M | 376 | 78 | 4.821 | 23 | 208 | 63 | 14 | 2 | 4 | 4 | 8 | 2 | ||
| D. scabra | MSNG 29239 | SOMALIA | Btwn Comia and Matagoi | M | 462 | 74 | 6.243 | 25 | 214 | 62 | 13 | 2 | 4 | 4 | 6 | 2 | 1 | |
| D. scabra | MZUF 31459* | SOMALIA | Ceelcillan = El Ellan | M | 389 | 81 | 4.802 | 25 | 204 | 62 | 14 | 2 | 4 | 4 | 6 | 74 | 2 | 0 |
| D. scabra | MZUF 24674* | SOMALIA | Dinsoor | M | 285 | 58 | 4.914 | 24 | 209 | 64 | 14 | 2 | 4 | 4 | 6 | 70 | 2 | 2 |
| D. scabra | MSNM 2071 | SOMALIA | Giohar | M | 397 | 22 | 203 | 14 | 2 | 4 | 4 | 4 | 50 | |||||
| D. scabra | MZUF 21968* | SOMALIA | Lugh – Ganame | M | 391 | 78 | 5.013 | 24 | 210 | 62 | 14 | 2 | 4 | 4 | 6 | 76 | 2 | 1 |
| D. scabra | MZUF 1134 | SOMALIA | Maheddei Uen | M | 461 | 89 | 5.18 | 22 | 209 | 63 | 14 | 2 | 4 | 4 | 6 | 2 | 2 | |
| D. scabra | FMNH 134939* | SOUTH SUDAN | Melut | M | 370 | 72 | 5.139 | 25 | 213 | 62 | 13 | 2 | 4 | 4 | 6 | 64 | 1 | 0 |
| D. scabra | NMW 9964/6+ | SOUTH SUDAN | ‘Sudan’ (taken as East Equatoria) | M | 449 | 88 | 5.102 | 23 | 215 | 67 | 14 | 2 | 4 | 4 | 4 | 59 | 1 | 0 |
| D. scabra | AMNH 50791 | TANZANIA | Chai Camp | M | 241 | 45 | 5.356 | 27 | 208 | 69 | 14 | 2 | 4 | 4 | 8 | 2 | ||
| D. scabra | NMW 9964/17 | TANZANIA | Dar es Salaam | M | 23 | 205 | 14 | 2 | 4 | 4 | 6 | 60 | 1 | |||||
| D. scabra | ZMB 26832* | TANZANIA | Dar es Salaam | M | 369 | 82 | 4.5 | 24 | 201 | 68 | 14 | 2 | 0 | 4 | 5 | 69 | 2 | 0 |
| D. scabra | BM 1954.1.2.154* | TANZANIA | Kange Estate | M | 400 | 85 | 4.706 | 23 | 206 | 67 | 14 | 2 | 2 | 4 | 6 | 71 | 2 | 1 |
| D. scabra | ZMB 22754 | TANZANIA | Kibwesa | M | 432 | 77 | 5.61 | 25 | 221 | 61 | 14 | 2 | 4 | 4 | 7 | 1 | 0 | |
| D. scabra | MCZ 30205 | TANZANIA | Maji Malulu, Usandawi | M | 181 | 27 | 225 | 14 | 2 | 4 | 4 | 6 | 54 | 2 | 1 | |||
| D. scabra | ZMB 14097* | TANZANIA | Massailand | M | 387 | 78 | 4.962 | 27 | 223 | 69 | 14 | 2 | 3 | 4 | 6 | 67 | 1 | 0 |
| D. scabra | USNM 62902* | TANZANIA | Morogoro | M | 335 | 64 | 5.234 | 24 | 218 | 62 | 14 | 3 | 4 | 4 | 5 | 69 | 2 | 0 |
| D. scabra | ZMB 19669* | TANZANIA | Mpwapwa | M | 398 | 87 | 4.575 | 26 | 214 | 69 | 14 | 2 | 4 | 4 | 6 | 54 | 2 | 0 |
| D. scabra | NMK 1887 | TANZANIA | Seronera, Serengeti N.P. | M | 344 | 73 | 4.712 | 211 | 72 | 14 | 4 | |||||||
| D. scabra | ZMB 16753* | TANZANIA | Wembere Steppe | M | 418 | 83 | 5.036 | 26 | 215 | 63 | 14 | 2 | 4 | 4 | 7 | 56 | 2 | 1 |
| D. scabra | BM 68.2.29.101* | TANZANIA | Zanzibar | M | 370 | 74 | 5 | 24 | 214 | 72 | 12 | 2 | 4 | 4 | 6 | 47 | 2 | 0 |
| D. scabra | MCZ 18208* | TANZANIA | Zengeragusa, Mkalama | M | 460 | 78 | 5.897 | 25 | 226 | 58 | 14 | 2 | 4 | 4 | 6 | 79 | 1 | 1 |
| D. scabra | LIV 1962.330.22 | UGANDA | Kayanja, Nyabusonji | M | 360 | 76 | 4.737 | 200 | 66 | |||||||||
| D. scabra | BM 51.1.5.79* | UGANDA | Mbarara, Ankole | M | 375 | 84 | 4.464 | 25 | 207 | 68 | 14 | 2 | 4 | 4 | 7 | 78 | 2 | 0 |
| D. scabra | LIV 1960.67.4 | UGANDA | Ruborogoto, Ishingiro | M | 198 | 38 | 5.211 | 188 | 63 | 4 | 4 | 6 | ||||||
| D. scabra | LIV 1960.67.7 | UGANDA | Ruborogoto, Ishingiro | M | 329 | 72 | 4.569 | 200 | 65 | 4 | 4 | 8 | ||||||
| D. scabra | LIV 1962.168.14 | UGANDA | Ruzinga, Ankole | M | 453 | 98 | 4.622 | 199 | 63 | 4 | 4 | 5 | ||||||
| D. scabra | LIV 1960.67.8 | UGANDA | Rwensama, Kashali | M | 352 | 83 | 4.241 | 185 | 64 | 4 | 4 | 6 | ||||||
| D. scabra | BM 07.12.1.19* | UGANDA | Wadelai | M | 333 | 67 | 4.97 | 23 | 214 | 67 | 14 | 2 | 4 | 4 | 6 | 65 | 1 | 0 |
| D. scabra | MNHN 6565* | ‘ARABIA’ | ? | F | 252 | 39 | 6.462 | 24 | 244 | 54 | 14 | 2 | 4 | 4 | 6 | 55 | 2 | 1 |
| D. scabra | MRAC 18563* | BURUNDI | Kabuyenge | F | 702 | 103 | 6.816 | 26 | 234 | 60 | 14 | 2 | 2 | 2 | 4 | 75 | 2 | 1 |
| D. scabra | MRAC 18565* | BURUNDI | Kahiro Dist. | F | 547 | 77 | 7.104 | 25 | 227 | 53 | 14 | 2 | 4 | 4 | 6 | 52 | 2 | 1 |
| D. scabra | MRAC 18564* | BURUNDI | Mugera | F | 495 | 72 | 6.875 | 25 | 220 | 52 | 14 | 2 | 4 | 4 | 6 | 60 | 2 | 1 |
| D. scabra | NMW 15298* | ETHIOPIA | ? | F | 590 | 87 | 6.782 | 28 | 219 | 53 | 14 | 2 | 4 | 6 | 7 | 56 | 2 | 1 |
| D. scabra | AAU 494 | ETHIOPIA | Arba Minch | F | 229 | 48 | 14 | 2 | 4 | 4 | 6 | |||||||
| D. scabra | AAU 679 | ETHIOPIA | Arba Minch | F | 668 | 92 | 7.261 | 22 | 243 | 54 | 14 | 2 | 4 | 4 | 5 | |||
| D. scabra | BM 16.6.24.12* | ETHIOPIA | Aware Melca | F | 415 | 71 | 5.845 | 24 | 221 | 59 | 14 | 2 | 4 | 4 | 6 | 56 | 1 | 0 |
| D. scabra | ZFMK 17763* | ETHIOPIA | Bulcha | F | 522 | 80 | 6.525 | 27 | 241 | 50 | 14 | 2 | 4 | 4 | 6 | 79 | 1 | 0 |
| D. scabra | PEM 8603* | ETHIOPIA | Entrance to Awash Nat. Park | F | 217 | 29 | 7.483 | 24 | 234 | 57 | 14 | 2 | 4 | 4 | 6 | 54 | 2 | 1 |
| D. scabra | BM 1964.1069 | ETHIOPIA | Gojam District | F | 525 | 76 | 6.908 | 23 | 219 | 47 | 12 | 2 | 4 | 4 | 7 | 0 | 0 | |
| D. scabra | ZMB 77053* | ETHIOPIA | Meki | F | 825 | 121 | 6.818 | 25 | 231 | 57 | 14 | 2 | 4 | 4 | 6 | 85 | 0 | 2 |
| D. scabra | BM 00.11.29.2* | ETHIOPIA | W of Omo River | F | 593 | 87 | 6.816 | 25 | 226 | 53 | 14 | 2 | 4 | 4 | 6 | 55 | 2 | 1 |
| D. scabra | PEM 5627 | KENYA | 45 km W of Ewaro Ngiro (Narok) | F | 26 | 236 | 49 | 14 | 2 | 4 | 4 | 7 | 1 | 0 | ||||
| D. scabra | USNM 42016 | KENYA | Aberdare Mtns | F | 305 | 38 | 8.026 | 26 | 220 | 47 | 14 | 2 | 4 | 4 | 6 | 69 | ||
| D. scabra | NMK 2824 | KENYA | Buna, Wajir Dist. | F | 411 | 73 | 5.63 | 222 | 55 | 14 | 4 | 67 | ||||||
| D. scabra | MSNPV CR791* | KENYA | Gatab, Mount Kulal | F | 324 | 61 | 5.311 | 22 | 219 | 64 | 14 | 2 | 4 | 4 | 6 | 94 | 1 | 1 |
| D. scabra | BM 01.1.3.4* | KENYA | Gilgil | F | 532 | 60 | 8.867 | 28 | 229 | 43 | 14 | 4 | 4 | 5 | 6 | 73 | 2 | 0 |
| D. scabra | CAS 147836 | KENYA | Hell’s Gate, Naivasha | F | 350 | 42 | 8.333 | 26 | 233 | 46 | 14 | 2 | 4 | 4 | 6 | 2 | 1 | |
| D. scabra | CAS 130784* | KENYA | Isiolo | F | 550 | 76 | 7.237 | 26 | 234 | 51 | 14 | 2 | 4 | 4 | 7 | 88 | 1 | 0 |
| D. scabra | BM 51-1-3-70 | KENYA | Kilifi | F | 480 | 25 | 227 | 14 | 2 | 2 | 4 | 6 | 70 | 2 | 0 | |||
| D. scabra | MCZ 40584* | KENYA | Laini, Tana River | F | 427 | 62 | 6.887 | 25 | 225 | 53 | 14 | 2 | 4 | 4 | 6 | 83 | 1 | 0 |
| D. scabra | NMK 1415 | KENYA | Lake Naivasha | F | 230 | 50 | 14 | 4 | 72 | 2 | 1 | |||||||
| D. scabra | NMK 2083 | KENYA | Lake Naivasha | F | 229 | 52 | 14 | 4 | 72 | 1 | 2 | |||||||
| D. scabra | NMK 2855 | KENYA | Lake Naivasha | F | 247 | 240 | 14 | 4 | 74 | |||||||||
| D. scabra | NMK 3184 | KENYA | Lake Naivasha | F | 867 | 230 | 14 | 4 | ||||||||||
| D. scabra | NMK 3319 | KENYA | Lake Naivasha | F | 657 | 98 | 6.704 | 215 | 50 | 14 | 4 | 72 | ||||||
| D. scabra | NMP 1260* | KENYA | Lake Naivasha | F | 509 | 76 | 6.697 | 27 | 228 | 52 | 14 | 2 | 4 | 4 | 8 | 72 | 1 | 1 |
| D. scabra | NMK 2819 | KENYA | Lolgorien, Trans-Mara | F | 372 | 58 | 6.414 | 232 | 55 | 14 | 6 | 69 | ||||||
| D. scabra | NMK 3005 | KENYA | Lolgorien, Trans-Mara | F | 281 | 42 | 6.69 | 233 | 53 | 14 | 4 | 73 | ||||||
| D. scabra | NMK 930 | KENYA | Manyani, Tsavo West N.P. | F | 368 | 57 | 6.456 | 231 | 52 | 14 | 4 | 60 | ||||||
| D. scabra | MCZ 40583 | KENYA | Mombosasa, near Witu | F | 648 | 97 | 6.68 | 25 | 228 | 55 | 14 | 2 | 4 | 4 | 6 | 76 | ||
| D. scabra | CAS 147837* | KENYA | Naivasha | F | 300 | 34 | 8.824 | 26 | 238 | 45 | 14 | 2 | 4 | 4 | 6 | 67 | 1 | 1 |
| D. scabra | CAS 147838* | KENYA | Naivasha | F | 325 | 40 | 8.125 | 25 | 226 | 45 | 14 | 2 | 4 | 4 | 6 | 75 | 2 | 1 |
| D. scabra | NMK 3103 | KENYA | Ol Ari Nyiro Ranch | F | 390 | 53 | 7.358 | 228 | 49 | 14 | 4 | 75 | ||||||
| D. scabra | USNM 41974 | KENYA | Sotik, Uaso Nyiro | F | 343 | 42 | 8.167 | 26 | 230 | 45 | 14 | 2 | 4 | 4 | 6 | 71 | 2 | |
| D. scabra | MCZ 11501* | KENYA | Uaso Nyiro Plains | F | 362 | 46 | 7.87 | 26 | 221 | 47 | 14 | 2 | 4 | 3 | 6 | 75 | 1 | 1 |
| D. scabra | CAS 130661 | KENYA | Wajir | F | 565 | 95 | 5.947 | 26 | 65 | 14 | 2 | 4 | 4 | 6 | 56 | 2 | 2 | |
| D. scabra | NMK 2262 | KENYA | Wajir | F | 232 | 50 | 14 | 2 | 63 | 2 | 0 | |||||||
| D. scabra | MCZ 40585* | KENYA | Wema (Ngatana) | F | 268 | 40 | 6.7 | 25 | 215 | 55 | 14 | 2 | 4 | 4 | 6 | 73 | 0 | 0 |
| D. scabra | MRAC 9214* | RWANDA | Gabiro | F | 637 | 83 | 7.675 | 25 | 235 | 48 | 14 | 2 | 4 | 5 | 6 | 83 | 2 | 0 |
| D. scabra | MZUF 1118* | SOMALIA | Afgoi | F | 180 | 31 | 5.806 | 25 | 218 | 56 | 14 | 2 | 4 | 6 | 7 | 55 | 0 | 0 |
| D. scabra | MZUF 1133* | SOMALIA | Afgoi | F | 344 | 57 | 6.035 | 24 | 228 | 64 | 14 | 2 | 4 | 4 | 7 | 74 | 2 | 1 |
| D. scabra | MSNG 42710A | SOMALIA | Belet Amin | F | 587 | 78 | 7.526 | 23 | 216 | 51 | 14 | 2 | 4 | 4 | 6 | 1 | 0 | |
| D. scabra | MZUF 24996* | SOMALIA | Dinsoor | F | 557 | 93 | 5.989 | 28 | 222 | 57 | 14 | 2 | 4 | 4 | 7 | 68 | 2 | 0 |
| D. scabra | MSNG 28808 | SOMALIA | Lugh | F | 519 | 66 | 7.864 | 24 | 222 | 54 | 14 | 4 | 4 | 6 | 4 | 2 | 1 | |
| D. scabra | BM 98.1.28.14* | SOMALIA | Lugh | F | 218 | 32 | 6.813 | 26 | 227 | 58 | 14 | 2 | 4 | 4 | 6 | 58 | 2 | 0 |
| D. scabra | MSNM 2148 | SOMALIA | Magadi (= Uagadi) | F | 496 | 80 | 6.2 | 22 | 202 | 57 | 14 | 2 | 4 | 4 | 6 | 51 | ||
| D. scabra | MZUF 2802* | SOMALIA | Mogadishu | F | 210 | 36 | 5.833 | 25 | 224 | 59 | 13 | 2 | 4 | 4 | 6 | 76 | 2 | 0 |
| D. scabra | MZUF 27156* | SOMALIA | near Mareri | F | 367 | 57 | 6.439 | 24 | 215 | 54 | 14 | 2 | 4 | 4 | 7 | 79 | 1 | 0 |
| D. scabra | RMNH 24399 | SUDAN | 40 km E of Rabak | F | 222 | 55 | 50 | |||||||||||
| D. scabra | MNHN 7227* | SUDAN | Nil Blanc | F | 673 | 90 | 7.478 | 25 | 226 | 52 | 14 | 2 | 4 | 4 | 6 | 55 | 1 | 1 |
| D. scabra | BM 09.10.15.35 | SUDAN | Sennar Province | F | 240 | 23 | 227 | 14 | 2 | 4 | 4 | 6 | 65 | 1 | 1 | |||
| D. scabra | ZMB 21062* | TANZANIA | Amani, Usambara Mtns | F | 740 | 124 | 5.968 | 27 | 234 | 65 | 14 | 2 | 4 | 4 | 7 | 63 | 2 | 1 |
| D. scabra | PEM 5626* | TANZANIA | Mapenduzi, Dodoma | F | 265 | 42 | 6.31 | 26 | 249 | 60 | 14 | 2 | 3 | 6 | 10 | 60 | 2 | 2 |
| D. scabra | ZMB 11934A* | TANZANIA | Massailand | F | 283 | 41 | 6.902 | 26 | 231 | 61 | 14 | 2 | 3 | 3 | 6 | 59 | 2 | 0 |
| D. scabra | ZMB 11934B* | TANZANIA | Massailand | F | 793 | 110 | 7.209 | 27 | 242 | 54 | 14 | 2 | 4 | 4 | 6 | 69 | 2 | 0 |
| D. scabra | USNM 62903 | TANZANIA | Morogoro | F | 500 | 85 | 5.882 | 26 | 232 | 57 | 14 | 2 | 4 | 5 | 8 | 67 | 0 | |
| D. scabra | USNM 62905 | TANZANIA | Morogoro | F | 447 | 72 | 6.208 | 26 | 223 | 59 | 14 | 2 | 4 | 4 | 6 | 79 | 2 | |
| D. scabra | BM 87.11.3.30* | TANZANIA | Mount Kilimanjaro | F | 346 | 52 | 6.654 | 25 | 230 | 58 | 14 | 2 | 2 | 3 | 6 | 67 | 1 | 0 |
| D. scabra | ZMB 13304* | TANZANIA | Mpwapwa | F | 403 | 63 | 6.397 | 27 | 225 | 57 | 14 | 2 | 2 | 4 | 6 | 62 | 2 | 0 |
| D. scabra | FMNH 27786 | TANZANIA | Ngare, Nanyuki | F | 539 | 26 | 229 | 55 | 14 | 2 | 4 | 4 | 8 | 69 | 2 | |||
| D. scabra | NMZB 1681* | TANZANIA | Tumba, Rukwa Valley | F | 710 | 103 | 6.893 | 26 | 238 | 54 | 14 | 2 | 3 | 4 | 6 | 60 | 2 | 2 |
| D. scabra | NMZB 659* | TANZANIA | Tumba, Rukwa Valley | F | 760 | 109 | 6.972 | 26 | 236 | 53 | 14 | 2 | 4 | 4 | 6 | 65 | 2 | 2 |
| D. scabra | MCZ 30207* | TANZANIA | Ukerewe Island | F | 524 | 70 | 7.486 | 26 | 223 | 49 | 14 | 2 | 4 | 4 | 8 | 62 | 2 | 1 |
| D. scabra | MCZ 30208 | TANZANIA | Ukerewe Island | F | 708 | 25 | 231 | 14 | 3 | 4 | 4 | 6 | 64 | 2 | 1 | |||
| D. scabra | BM 68.2.29.100* | TANZANIA | Zanzibar | F | 476 | 78 | 6.103 | 25 | 226 | 59 | 13 | 2 | 4 | 4 | 6 | 46 | 2 | 0 |
| D. scabra | BM 68.2.29.102* | TANZANIA | Zanzibar | F | 335 | 51 | 6.569 | 27 | 233 | 58 | 14 | 2 | 4 | 4 | 7 | 53 | 2 | 0 |
| D. scabra | LIV 1962.106.6 | UGANDA | Kaburangiri, Kashali | F | 270 | 32 | 8.438 | 216 | 52 | 4 | 4 | 5 | ||||||
| D. scabra | LIV 1960.67.6 | UGANDA | Kakoba, Kashari | F | 612 | 85 | 7.2 | 216 | 49 | 3 | 4 | 6 | ||||||
| D. scabra | LIV 1959.83.1 | UGANDA | Mbarara, Ankole | F | 640 | 90 | 7.111 | 217 | 51 | 2 | ||||||||
| D. scabra | BM 1961.1804* | UGANDA | Mbarara, Kashari | F | 690 | 105 | 6.571 | 25 | 216 | 53 | 14 | 2 | 4 | 4 | 6 | 71 | 2 | 2 |
| D. scabra | BM 03.12.2.21* | UGANDA | Mulema | F | 550 | 84 | 6.548 | 27 | 219 | 54 | 14 | 2 | 4 | 4 | 6 | 72 | 2 | 0 |
| D. scabra | LIV 1962.106.5 | UGANDA | Namatyobola, Kashali | F | 354 | 53 | 6.679 | 206 | 51 | 3 | 4 | 6 | ||||||
| D. scabra | LIV 1962.106.35 | UGANDA | Ruborogoto, Ishingiro | F | 810 | 113 | 7.168 | 222 | 53 | 4 | 4 | 6 | ||||||
| D. scabra | LIV 1962.106.20 | UGANDA | Rukoni, Kwampala | F | 480 | 63 | 7.619 | 220 | 50 | 4 | 4 | 6 | ||||||
| D. scabra | BM 1959.1.5.59 | UGANDA | Ruzinga, Isingiro | F | 650 | 92 | 7.065 | 230 | 57 | 14 | 2 | 4 | 4 | 6 | 78 | 1 | 1 | |
| D. scabra | BM 1969.288 | ETHIOPIA | Arba Minch | U | 155 | 27 | 5.741 | 26 | 213 | 59 | 14 | 2 | 4 | 4 | 7 | 82 | 0 | 0 |
| D. scabra | BM 16.6.24.11 | ETHIOPIA | Aware Melca | U | 210 | 28 | 7.5 | 25 | 235 | 55 | 14 | 2 | 4 | 4 | 6 | 66 | 1 | 1 |
| D. scabra | IRSNB 5499 | RWANDA | Gabiro | U | 443 | 66 | 6.712 | 26 | 226 | 54 | 14 | 2 | 4 | 4 | 5 | 70 | ||
| D. scabra | PEM 5625 | RWANDA | Nyagatare | U | 163 | 31.5 | 5.094 | 25 | 194 | 59 | 14 | 2 | 4 | 4 | 6 | 55 | 2 | 0 |
| D. scabra | USNM 72476 | TANZANIA | Kikombo, Dodoma | U | 200 | 28 | 7.143 | 26 | 253 | 67 | 14 | 2 | 4 | 4 | 6 | 68 | 1 | |
| D. scabra | ZMUC 6136 | TANZANIA | Minziro Forest Reserve | U | 176 | 27 | 6.519 | 25 | 216 | 52 | 11 | 2 | 4 | 3 | 5 | 63 | 2 | 1 |
| D. scabra | ZMUC 6137 | TANZANIA | Minziro Forest Reserve | U | 188 | 31 | 6.065 | 26 | 226 | 56 | 14 | 2 | 4 | 4 | 6 | 1 | 0 | |
| D. scabra | NMW 9964/9+ | TANZANIA | Tanga | U | 195 | 37 | 5.270 | 25 | 204 | 63 | 14 | 2 | 3 | 4 | 6 | 65 | 1 | 0 |
| D. taylori | BM 1992.501 | SOMALIA | 2.5 km SW of Hargeisa | M | 305 | 199 | 14 | 2 | 4 | 4 | 6 | 44 | 2 | 1 | ||||
| D. taylori | BM 51.1.7.69* | SOMALIA | Garadak | M | 23 | 202 | 59 | 14 | 2 | 4 | 4 | 4 | 49 | 2 | 1 | |||
| D. taylori | BM 49.2.2.1* | SOMALIA | Haud | M | 188 | 34 | 5.529 | 23 | 196 | 55 | 12 | 2 | 3 | 4 | 6 | 55 | 2 | 0 |
| D. taylori | BM 49.2.2.2* | SOMALIA | Haud | M | 172 | 30 | 5.733 | 24 | 198 | 60 | 14 | 2 | 4 | 4 | 6 | 50 | 2 | 1 |
| D. taylori | BM 49.2.2.5 | SOMALIA | Ogo | M | 408 | 83 | 4.916 | 25 | 216 | 68 | 14 | 2 | 4 | 4 | 6 | 53 | 0 | |
| D. taylori | MNHN 84-707* | DJIBOUTI | ? | F | 477 | 68 | 7 | 25 | 222 | 52 | 14 | 2 | 4 | 4 | 6 | 59 | 2 | 1 |
| D. taylori | BM 49.2.2.4 | SOMALIA | Hargeisa Dist. | F | 530 | 24 | 208 | 14 | 2 | 2 | 2 | 6 | 53 | 2 | 1 | |||
| D. taylori | BM 49.2.2.3* | SOMALIA | Haud | F | 189 | 23 | 8.217 | 24 | 211 | 44 | 14 | 2 | 3 | 4 | 6 | 59 | 2 | 1 |
FIGURE CAPTIONS

Figure 1. Tail length relative to snout-vent vent in males and females of Dasypeltis atra, D. confusa, D. fasciata, D. medici, D. scabra and D. abyssina in north-eastern Africa.

Figure 2. Dasypeltis medici. A: Vicinity of Dar-es-Salaam, Tanzania (Photo: W.R. Branch); B: Juvenile from East Usambara Mountains, Tanzania (Photo: W.R. Branch); C: Watamu, Kenya (Photo: S. Spawls); D: Tanga, Tanzania (Photo: S. Spawls); E & F: Diani Beach, Kenya (Photos: F. Finke). [A & B are colour pattern ‘8’ (‘medici’), C & D are pattern ‘3’ (‘lamuensis’, mostly unmarked), and E & F—same individual—are intermediate (weakly marked)]

Figure 3. Geographical distribution of the genus Dasypeltis in north-eastern Africa and south-western Arabia.

Figure 4. Dasypeltis confusa. [‘5L’ colour pattern of Gans 1959] A: Bisu (Kigezi), Uganda. (BM 1951.1.3.69, Photo: M.F. Bates); B: Kacheliba, Kenya (NMK 489, Photo: P. Malonza); C: Mumias, Kenya (CAS 141784, Photo: J. Vindum); D: Lake Kivu, Rwanda (ZMB 77441, Photo: F. Tillack).

Figure 5. Dasypeltis scabra. [‘5N’ colour pattern of Gans 1959] A: Dodoma, Tanzania (Photo: W.R. Branch); B: Masiko Tembo, Tanzania (Photo: W.R. Branch), C: Wajir, Kenya (Photo: S. Spawls); D: Juvenile from Loita Hills, Kenya (Photo: T. Mazuch); E: Gilgil, Kenya (Photo: S. Spawls); F: MSNM 974, Magadi, Somalia (Photo: Museo Civico di Storia Naturale, Milan); G: Awash National Park, Ethiopia (Photo: S. Spawls); H: Awash National Park, Ethiopia (Photo: S. Spawls).

Figure 6. Dasypeltis scabra. Close-up dorsal view (in alcohol) of MNHN 7227 (syntype of Rachiodon abyssinus) from the White Nile River (possibly near Khartoum, Sudan) showing a ‘5N’ pattern (see Gans 1959) consisting of dark oval saddles and dark dorso-ventrally elongated lateral bars, with paler centres. (Photo: M.F. Bates)

Figure 7. Dasypeltis bazi. A: Dorsal and lateral views of the largest known female (BM 97.10.28.575) from Fayoum, Egypt (Photo: M.F. Bates); B: Ventral view of the same specimen (Photo: Natural History Museum, London).
Figure 8. Dasypeltis bazi. Dorsal (left), lateral (middle) and ventral (right) views of the head of the largest known female (BM 97.10.28.575) from Fayoum, Egypt. (Photo: Natural History Museum, London)

Figure 9. Dasypeltis bazi. Dorsal (A) and ventral (B) views of a male (ZMB 1861) from ‘Upper Egypt’. (Photo: F. Tillack)

Figure 10. Dasypeltis bazi. Dorsal (left), lateral (middle) and ventral (right) views of the head of a male (ZMB 1861) from ‘Upper Egypt’. (Photo: F. Tillack)

Figure 11. Dorsal views of Dasypeltis loveridgei comb. nov. [‘6N’ colour pattern of Gans 1959; specimen also has 6 supralabials on either side of head, with 2nd and 3rd in contact with orbital] (female, ZMB 33670a, top) from Otjimbingwe, Namibia, and Dasypeltis bazi (male, ZMB 1861, below) from ‘Upper Egypt’ showing the general similarity in colour pattern. (Photo: M.F. Bates)

Figure 12. Dasypeltis cf. bazi. Dorsal view of the anterior part of the body of a specimen (ZFMK 38415) from Erkowit, Sudan. (Photo: W. Böhme)

Figure 13. Dasypeltis taylori sp. nov. Body and head views of the holotype (BM 1949.2.2.2) from Haud, Somaliland. (Photos: Natural History Museum, London)

Figure 14. Dasypeltis taylori sp. nov. Paratype (MNHN 1884.0707) from Djibouti. (Photo: I. Ineich)

Figure 15. Haud, Somaliland—type locality of Dasypeltis taylori sp. nov. (Photo: T. Mazuch)

Figure 16. Vicinity of Hargeisa, Somaliland—habitat of Dasypeltis taylori sp. nov. (Photo: T. Mazuch)

Figure 17. Dasypeltis atra. Black morph (‘2M’ of Gans 1959)—A: Keriyo near Mulu Cattle Farm, Ethiopia (Photo: S. Spawls); B: Limuru, Kenya (Photo: S. Spawls); Brown morph (‘2B’ of Gans 1959)—C: ELI 1264 (field number), Kibira Forest, Burundi (Photo: E. Greenbaum); Patterned morph (‘9’ of Gans 1959)—D: Holotype (ZMB 9224) of Dasypeltis lineolata Peters from Kitui, Kenya (Photo: M.F. Bates); Patterned morph (‘2Bx’ of Gans 1959)—E: Addis Ababa, Ethiopia (Photo: S. Spawls); F: Kitengela, Kenya (Photo: S. Spawls); G: Chuka Mountain, Kenya (Photo: S. Spawls); H: ELI 2760 (field number), Bwindi Impenetrable National Park, Uganda (Photo: E. Greenbaum); I: Addis Ababa, Ethiopia (Photo: S. Spawls); J: Masiko Tembo, Tanzania (Photo: W.R. Branch); K: PEM R17336, Klein’s Camp, Mkomazi Game Reserve, Tanzania (Photo: W.R. Branch); L: Kakamega Forest, Kenya (Photo: T. Mazuch); M: Neghelle, Ethiopia (Photo: V. Trailin).

Figure 18. Dasypeltis atra from Kakamega Forest, Kenya, with a single postocular. (Photo: T. Mazuch)

Figure 19. Dasypeltis fasciata. [Colour pattern ‘4’ (checker board) of Gans 1959] A & B: Dorsal and ventral views, respectively, of TCWC 23839, Nandi, Bwamba Forest, Uganda (Photos: T. Hibbits); C: Makeni, Sierra Leone (Photo: W.R. Branch).

Figure 20. Dasypeltis abyssina. Dorsal (A) and ventral (B) aspects of the lectotype (MNHN 6567) from Ethiopia (Photos: M.F. Bates); C: Mulu Cattle farm, Ethiopia (Photo: S. Spawls); D: Ethiopia (Photo: C. Tilbury); E: Mulu Cattle farm, Ethiopia (Photo: S. Spawls); F: Keriyo near Mulu, Ethiopia (Photo: S. Spawls); G: Eritrea (NMW 15301–3, Photo: M.F. Bates).

Figure 21. Habitat of Dasypeltis abyssina and D. atra near Lake Tana, north of Addis Zemen, Ethiopia. (Photo: T. Mazuch)

Figure 22. Habitat of Dasypeltis abyssina south of Gondar, northern Ethiopia. (Photo: T. Mazuch)

Figure 23. Dasypeltis crucifera sp. nov. Dorsal (A) and ventral (B) views of the holotype (ZMB 7631) from Bogos, Eritrea (Photo: F. Tillack).

Figure 24. Dasypeltis crucifera sp. nov. Dorsal (A), lateral (B) and ventral (C) aspects of the head of the holotype (ZMB 7631) from Bogos, Eritrea (Photo: F. Tillack).

Figure 25. Dasypeltis crucifera sp. nov. Dorsal (A) and ventral (B) views of the allotype (MZUT 3582) from Agordat, Eritrea (Photo: F. Andreone).

Figure 26. Keren, Eritrea—type locality of Dasypeltis crucifera sp. nov. (Photo: T. Mazuch)

Figure 27. Dasypeltis arabica sp. nov. [‘5I’ colour pattern of Gans 1959] Dorsal (A), ventral (B) and lateral (C) views of the holotype (BM 1987.2192) from Sana’a, northern Yemen. (Photos: M.F. Bates)
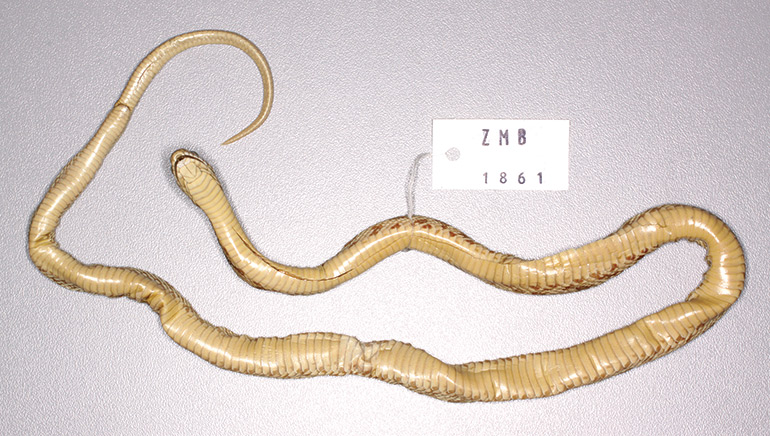
Figure 28. Dasypeltis arabica sp. nov. [‘5I’ colour pattern of Gans 1959] A: Dorsal (left), lateral (middle) and ventral (right) aspects of the head of the holotype (BM 1987.2192) from Sana’a, northern Yemen (Photo: Natural History Museum, London); B: Dorso-lateral aspect of the head of the holotype (in water, Photo: M.F. Bates).

Figure 29. Dasypeltis arabica sp. nov. [‘5I’ colour pattern of Gans 1959] A: Dorsal view of a paratype (BM 1987.1018) from Ajibah, Saudi Arabia. (Photo: M.F. Bates); B: Dorsal (left), lateral (middle) and ventral (right) aspects of the head of the paratype (Natural History Museum, London).
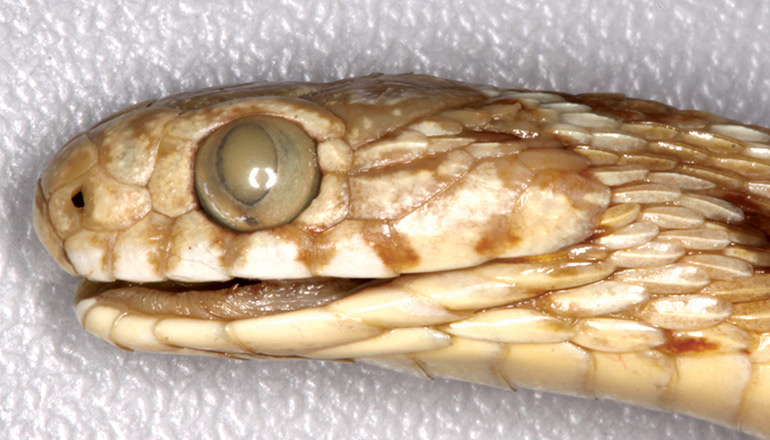
Figure 30. Echis megalocephalus (BM 1987.3217) from Nocra Island, Dahlak Archipelago, Eritrea—a possible model species for mimicry by Dasypeltis arabica sp. nov. and Dasypeltis crucifera sp. nov. (Photo: T. Mazuch)
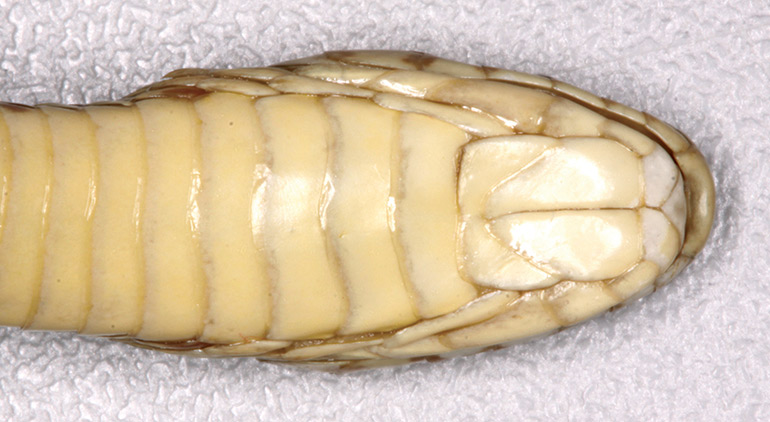
Figure 31. Principal Component Analysis of the genus Dasypeltis in north-eastern Africa and south-western Arabia. (MBSR = midbody scale rows, V = ventrals, SC = subcaudals, SL = supralabials, Pre Oc = preoculars, Pos Oc = postoculars, Ant T = anterior temporals, Pos T = posterior temporals, Cycles = pattern cycles, Frontal = frontal pitting, Sulcus = Inter-prefrontal sulcus)

Figure 32. Discriminant Function Analysis of the genus Dasypeltis in north-eastern Africa and south-western Arabia.

Figure 33. Principal Component Analysis of the genus Dasypeltis in the Horn of Africa and Egypt. (MBSR = midbody scale rows, V = ventrals, SC = subcaudals, SL = supralabials, Pre Oc = preoculars, Pos Oc = postoculars, Ant T = anterior temporals, Pos T = posterior temporals, Cycles = pattern cycles, Frontal = frontal pitting, Sulcus = Inter-prefrontal sulcus)

Figure 34. Discriminant Function Analysis of the genus Dasypeltis in the Horn of Africa and Egypt.



Comments are closed.