
J.A. NEETHLING1, 2 & C.R. HADDAD1
1Department of Zoology & Entomology, University of the Free State, P.O. Box 339, Bloemfontein, 9300 , South Africa
2Department of Arachnology, National Museum, P.O. Box 266, Bloemfontein 9300, South Africa. E-mail: ja.neethling@nasmus.co.za
ABSTRACT
The pseudoscorpion family Geogarypidae of South Africa was revised. Prior to this study the family consisted of eight species in two genera (Afrogarypus Beier, 1931 and Geogarypus Chamberlin, 1930). This study is the first holistic approach to the classification of South African pseudoscorpions, taking both morphological as well as molecular phylogenetic (COI and 28S ) data into consideration. Results showed that morphology and phylogenetics complemented each other and that there are 18 distinct species of Geogarypidae in South Africa, nine of which are new, namely A. carmenae sp. nov., A. castigatus sp. nov., A. megamolaris sp. nov., G. deceptor sp. nov., G. liomendontus sp. nov., G. modjadji sp. nov., G. octoramosus sp. nov., G. tectomaculatus sp. nov. and G. variaspinosus sp. nov. These are divided into three major clades, one corresponding to Geogarypus and two to Afrogarypus. Four species previously described in Geogarypus were found to belong to the Afrogarypus clades and are transferred: A. minutus (Tullgren, 1907) comb. nov., A. purcelli (Ellingsen, 1912) comb. nov., A. robustus (Beier, 1947) comb. nov. and A. triangularis (Ellingsen, 1912) comb. nov. The data also supports the separation of G. olivaceus (Tullgren, 1907) and G. flavus (Beier, 1947) (synonymised by Beier in 1964), with the latter re-validated. Lastly, A. excelsus excellens (Beier, 1964) was found to be a synonym of A. excelsus (Beier, 1964) stat. nov.
Key words: morphology, phylogenetics, geographical distribution, false-scorpions
CONTENTS
INTRODUCTION ………………………………………………………………………x
MATERIALS AND METHODS ………………………………………………………x
Specimen sampling ……………………………………………………………… x
Morphological analysis ……………………………………………………… x
Molecular methods ……………………………………………………………… x
SYSTEMATICS OF THE GEOGARYPIDAE ……………………………………… x
Key to the Geogarypidae genera and species of South Africa ………………. x
Afrogarypus carmenae sp. nov. …………………………………………………………. x
Afrogarypus castigatus sp. nov. …………………………………………………………. x
Afrogarypus excelsus (Beier, 1964) stat. nov. ……………………………………. x
Afrogarypus impressus (Tullgren, 1907) ………………………………………………. x
Afrogarypus megamolaris sp. nov. …………………………………………………………. x
Afrogarypus minutus (Tullgren, 1907) comb. nov. …………………………… x
Afrogarypus purcelli (Ellingsen, 1912) comb. nov. …………………………… x
Afrogarypus robustus (Beier, 1947) comb. nov. …………………………… x
Afrogarypus subimpressus (Beier, 1955) ………………………………………… x
Afrogarypus triangularis (Ellingsen, 1912) comb. nov. …………………… x
Geogarypus deceptor sp. nov. …………………………………………………………. x
Geogarypus flavus (Beier, 1947) stat. nov. ………………………………………………. x
Geogarypus liomendontus sp. nov. …………………………………………………………. x
Geogarypus modjadji sp. nov. …………………………………………………………. x
Geogarypus octoramosus sp. nov. …………………………………………………………. x
Geogarypus olivaceus (Tullgren, 1907) …………………………………… x
Geogarypus tectomaculatus sp. nov. …………………………………………………………. x
Geogarypus variaspinosus sp. nov. …………………………………………………………. x
BIOGEOGRAPHY …………………………………………………………………………………………….. x
SOUTH AFRICAN GEOGARYPIDAE PHYLOGENETICS …………………………….. x
DISCUSSION ……………………………………………………………………………………………………….. x
CONCLUSION …………………………………………………………………………………………….. x
ACKNOWLEDGEMENTS ………………………………………………………………………………….. x
REFERENCES …………………………………………………………………………………………….. x
Introduction
At the start of the 20th century data on pseudoscorpions from southern Africa was sparse at best, with Cordylochernes octentoctus (Balzan, 1892), Withius simoni (Balzan, 1892) and W. tenuimanus (Balzan, 1892) being the only South African endemics known (Ellingsen 1912). Historically, most of the early research, specifically species descriptions, was done by foreign scientists. The works of Tullgren (1907a, b), Ellingsen (1912), and to a lesser extent, Hewitt & Godfrey (1929), saw an abundance of new species descriptions. However, it was Max Beier who was by far the greatest contributor to the field, describing from 1947 to 1966 (Beier 1947, 1955, 1964, 1966), approximately 80% of the currently known species in the region. Local contributions were made by Lawrence (1935, 1967).
After 1964 the discovery of new species decreased drastically, with the last South African species described by Mahnert (1988). Dippenaar-Schoeman & Harvey (2000) published a complete checklist and catalogue of species found in South Africa, and in subsequent years several checklists of nature reserves included pseudoscorpions (Haddad et al. 2006; Haddad & Dippenaar-Schoeman 2009, 2015). In recent years South African specimens also contributed to karyotype studies (Šťáhlavský et al. 2006, 2012), as well as phylogenetic analyses (Murienne, Harvey & Giribet 2008; Van Heerden, Taylor & Van Heerden 2013).
South Africa currently has 152 known species in 17 families, with over 70% of these species endemic to the country, ranking the region eighth in the world with regards to pseudoscorpion species richness (Dippenaar-Schoeman & Harvey 2000; Harvey 2013). Since the last species description by Manhert (1988), the main focus of research has shifted to sampling and the creation of species checklists of the region. Following advances in pseudoscorpion taxonomy, both morphologically (Harvey 1992) as well as phylogenetically (Murienne et al. 2008), an opportunity presented itself to follow on their success and start revisions of the South African fauna.
The family Geogarypidae was chosen to spearhead this endeavour, as it had been revised by Harvey (1986) in Australia. The South African fauna consists of nine taxa in two genera (Afrogarypus Beier, 1931 and Geogarypus Chamberlin, 1930), with one species, A. excelsus (Beier, 1964), consisting of two subspecies. Difficulty soon arose in the acquisition of the type specimens. Since the indigenous fauna was described over a 60 year period by three foreign authors, many types were not lodged in South African collections. Although the types for G. flavus (Beier, 1947), G. minutus (Tullgren, 1907), G. olivaceus (Tullgren, 1907) and G. robustus (Beier, 1947) were located, the remaining types could not be found and are presumed to be lost or destroyed during World War II. A few are lodged in collections overseas, and since the curators were understandably unwilling to ship such valuable material, not all of the exsisting types could be examined, although curators kindly provided high resolution images of some types for comparison with recently collected material.
Fossil Geogarypidae bearing all the characteristic morphological traits of their modern descendents have been collected from both Baltic (Henderickx 2005) and Rovno (Henderickx & Perkovsky 2012) ambers dating from the late Eocene to early Oligocene epochs, strongly suggesting that members of this family have been around for at least 50 million years. Most modern species are generally tropical, subtropical or temperate, distributed north and south throughout the equatorial belt (Harvey 2013), and can commonly be found in leaf litter and under stones in habitats ranging in elevation from 0 – 3200m above sea level (Beron 2002). Most species are regional endemics with small distribution ranges, but due to human traffic, species such as Geogarypus mirei Heurtault, 1970 have extended their range beyond their natural dispersal capabilities (Mahnert 2011). Historically, most research focused on species descriptions, but pseudoscorpions have recently been investigated as potential biological control agents in certain crops (Devasahayam & Koya 1994).
Originally a subfamily of the Garypidae, Geogarypinae was elevated to full familial status by Harvey (1986). Previously all species were placed in the genus Geogarypus, which had been divided into the three subgenera: Geogarypus, Afrogarypus and Indogarypus by Beier (1931, 1947, 1955, 1957, 1964). Afrogarypus and Indogarypus were subsequently elevated to genus level by Harvey (1986) The family is currently represented by 67 species worldwide, of which 23 species have been described from the Afrotropical Region, five in Geogarypus and 18 in Afrogarypus.
While Indogarypus is restricted to India and Sri Lanka (Harvey 1986), both Afrogarypus and Geogarypus have much wider distributions, with both genera being represented by multiple species in South Africa (Tullgren 1907a, b; Ellingsen 1912; Beier 1947, 1955, 1964). There are currently eight recognized species and two subspecies in South Africa, namely, Afrogarypus excelsus excelsus; A. excelsus excellens; A. impressus; A. subimpressus; Geogarypus minutus; G. olivaceus; G. purcelli; G. robustus and G. triangularis.
This study aims to provide a holistic view of the taxonomy of the Geogarypidae of South Africa through the incorporation of both morphological and molecular phylogenetic characters in species descriptions. Keys to the species are also provided
Materials and methods
Specimen sampling
Between September 2011 and December 2013 a total of 53 locations were sampled across eight of the nine provinces of South Africa. Due to habitat homologies with much of the Free State and Northern Cape (Mucina & Rutherford 2006), the North West Province was not sampled. A single geogarypid museum sample from the North West Province was studied, and future work will include sampling in this province to fill the information gap.
Most type localities are situated within the Afromontane belt and coastal fynbos areas (Tullgren 1907a, b; Ellingsen 1912; Beier 1947, 1955, 1964), possibly due to sampling efforts that were more focused on coastal regions in the past. Effort was made to sample as many diverse habitats as possible in South Africa.
Leaf litter sifting
Sifting of leaf litter was done using a sieve containing a metal mesh with 5 mm spacing. The technique consisted of collecting leaf litter, as well as other detritus material, from the environment and sifting the material over a white sheet. Any pseudoscorpions could then be easily spotted, collected and placed in 1.5 ml cryovials with 99% ethanol for storage.
Canopy fogging
Fogging of the canopies of trees located at most of the sampling locations was conducted using a Stihl SR430 blower. A solution of Tobaccoguard®, a pyrethroid-based ULV spray, and D.B.M.® Double Strength, an organophosphate based emulsifiable concentrate, mixed in a ratio of 100:1, acted as the knock-down agent. White sheets mounted and spread on metal rods were placed under the target trees and covered an area of 54 m2 beneath the canopies. A total of one hour was allowed for the knockdown agent to work on each tree sampled, whereafter specimens were collected from the sheets using household Electrolux hand-held vacuum cleaners, fitted internally with a fine material mesh to stop any specimens from entering the motor and blades. Material was then emptied into 250 ml plastic bottles and filled with 99% ethanol. The bottles were later emptied into white plastic sorting trays in the laboratory and pseudoscorpions were collected and separated by morphospecies into 1.5 ml cryovials containing fresh 99% ethanol.
Hand collecting
Hand collecting consisted of lifting stones, logs and other debris in search of specimens. Occasionally shrubs were also sampled by beating vegetation using a 35 cm diameter sweep net and beating stick.
All specimen vials collected for an area were provided with detailed labels containing locality and sampling data. Locality co-ordinates were obtained using a Garmin GPSmap 62 handheld Global Positioning System (accuracy of 3–6 m). Specimens were deposited in the Arachnology collection of the National Museum, Bloemfontein (NMBA).
Morphological analysis
Geogarypidae specimens collected during field work were firstly separated into morphospecies. Using specimens from type localities, as well as available literature (Tullgren 1907a, b; Ellingsen 1912; Beier 1947, 1955, 1964), the nine described species were identified and also compared to all known Afrotropical Geogarypidae. All specimens not matching known species descriptions were initially identified to genus level, and designated as new species only after both morphological and molecular analysis. Additional specimens were either loaned from, or examined at, their respective institutions to aid in this study.
The following institutions either supplied, or currently house, specimens referred to in this study (curators in parentheses):
AMG Albany Museum, Grahamstown, South Africa (John Midgley)
NCA National Collection of Arachnida, ARC – Plant Protection Research Institute,
Pretoria, South Africa (Ansie Dippenaar-Schoeman, Petro Marais)
NMBA National Museum, Bloemfontein, South Africa (Leon Lotz)
NMSA KwaZulu-Natal Museum, Pietermaritzburg, South Africa (Burgert Muller)
NMZA Natural History Museum of Zimbabwe, Bulawayo, Zimbabwe (Moira FitzPatrick)
SAMC Iziko South African Museum, Cape Town, South Africa (Dawn Larsen)
WAM Western Australian Museum, Perth, Australia (Mark Harvey)
ZMH Zoological Museum of Hamburg, Hamburg, Germany (Markus Koch)
During discussions with the Ditsong Museum, Gauteng, it was established that no pseudoscorpion specimens belonging to the Geogarypidae could be located. Most of the slides from AMG were damaged during a fire in 1941, which resulted in poor specimen visibility. Although specimens could be identified, the sexes of most could not be determined. Where locality co-ordinates could not be located either on labels or in an institution’s database, they were estimated by georeferencing, mainly using the Global Gazetteer version 2.2 (http://www.fallingrain.com) and Google Earth (https://www.google.com/earth/). Such localities are indicated by square brackets in the lists of material examined. Distribution maps were generated using the software program Quantum GIS Brighton version 2.6.1 (http://www.qgis.org/en/site/).
Morphological techniques used:
Lactic acid clearing
Before clearing, male and female representatives of each species were transferred into glycerin and left overnight. This step prevented exemplars from floating to the top during the clearing step, resulting in uneven to no clearing. Specimens were then transferred into small glass vials containing 90% lactic acid and left overnight to clear. For some heavily sclerotised specimens, such as those of Afrogarypus impressus (Tullgren, 1907), it was necessary to clear the specimens for two days. After clearing, specimens were dissected according to Hu & Zhang (2012) by removing the chelicerae, pedipalps, leg I and leg IV using size 0 insect pins. The individual structures were then temporarily mounted in the same lactic acid on standard microscope slides. Following Harvey (2010), small pieces of nylon fishing line were used to elevate the cover slip above the structures. This prevented the latter from being crushed, while providing room for the rotation of the structures by movement of the cover slip. Fishing line thicknesses used were: 0.12 mm for chelicerae, 0.26 mm for pedipalps and legs, and 0.45 mm for carapace and abdomen. Slides were then mounted on an Axiophot stereoscopic microscope (Zeiss, Germany) fitted with an AxioCam ICc 5. Images and measurements were done using the 64bit software AxioVision Special Edition version 4.9.1 (http://www.zeiss.co.za/microscopy/en_za/home.html). Extended focal range images of studied structures were produced by stacking in Adobe Photoshop CS5. These images were then printed and traced to produce line drawings used for species descriptions.
Scanning electron microscopy (S.E.M.)
Specimens were cleaned of soil and other debris using a Branson® 3200 ultrasonic bath at 30 s intervals for 3 min while suspended in 10 ml glass vials containing 100% ethanol. Images were produced on a Shimadzu SSX-550 S.E.M. (Kyoto, Japan). Specimens were first dehydrated using 100% ethanol and then critical-point-dried using a Tousimis critical point dryer (Rockville, Maryland, U.S.A.) and carbon dioxide drying gas. After being mounted on stubs using thin double-sided tape, specimens were gold-sputter-coated at 50–60 nm thickness in a BIO-RAD (Microscience division) coating system (London, U.K.). All images were produced at 5.00 kV. All stubs were donated to the Centre for Microscopy at the University of the Free State on their request.
Terminology mostly follows Harvey (1992) with the exception of the chelicerae, which follows Judson (2007). Ratios are given as length/width. The following abbreviations are used in the figures and text:
Chela trichobothria Chelicerae
| b | = | Basal | es | = | exterior seta |
| sb | = | sub-basal | bs | = | basal seta |
| st | = | sub-terminal | sbs | = | sub-basal seta |
| t | = | Terminal | is | = | interior seta |
| ib | = | interior basal | ls | = | laminal seta |
| isb | = | interior sub-basal | gs | = | galea seta |
| ist | = | interior sub-terminal | se | = | serula exterior |
| it | = | interior terminal | si | = | serula interior |
| eb | = | exterior basal | le | = | lamina exterior |
| esb | = | exterior sub-basal | r | = | rallum |
| est | = | exterior sub-terminal | g | = | galea |
| et | = | exterior terminal |
Molecular methods
Using Murienne, Harvey & Giribet (2008) as a reference guide, the following steps were followed to determine the phylogenetic relationships between South African species of Geogarypidae using molecular techniques. The above-mentioned paper used three molecular markers consisting of two nuclear ribosomal genes (complete 1.8 kb 18S rRNA and a 1kb fragment of 28S rRNA) and one mitochondrial protein-coding gene (cytochrome c oxidase subunit one).
In the present study 18S rRNA was discarded due to its potential for weak resolution at species level branches (Steiner & Müller 1996), and the fact that it has to be sequenced in three fragments of 900 bp each, which was restricted due to funding limitations. It was therefore decided to rather focus on the remaining two genes, namely the nuclear gene 28S rRNA (28S hereafter) and the mitochondrial gene cytochrome c oxidase subunit one (COI hereafter).
One female specimen per morphospecies was designated for genetic analysis and stored in 1.5 ml cryovials in 100% ethanol at -80oC until DNA extraction. After DNA extraction the genomic DNA of each specimen was again stored at -80oC until amplification. All Polymerase Chain Reaction (PCR) products were stored at -20oC until sent for sequencing.
DNA extraction, amplification and sequencing
Initial DNA extraction was performed at the WAM (Perth, Australia) using a modified version of the salting out procedure of Miller, Dykes & Polesky (1988), as this was the standard procedure used at the facility. Unfortunately, not only was the procedure a time-consuming endeavour, taking three days to yield de-proteinised genomic DNA, but it failed to yield any genomic DNA in over 50% of samples.
Final DNA extraction was then performed by using the same method as Murienne et al. (2008). Tissue lysis and DNA purification was done using a DNEasy® tissue kit (Qiagen, R.S.A.), following the manufacturer’s protocol. Total genomic DNA was extracted by incubating crushed, whole specimens in the lysis buffer overnight. The purified genomic DNA was then used as a template for PCR amplification.
Primer-pairs used in this study:
| Gene | Primers | Sequence | Reference
|
| 28S | 28SpsF1 | 5′- ATTA CCC GCC GAA TTT AAGC -3′ | Murienne et al. (2008) |
| 28SpsR1 | 5′- TCG GAG GGA ACC AGC TAC -3′
|
||
| COI | LCO1490 | 5′- GGTC AAC AAA TCA TAA AGA TAT TGG -3′ | Folmer et al. (1994) |
| HCO2198 | 5′- TAAA CTT CAG GGT GAC CAA AAA ATCA -3′ |
The above primers resulted in the amplification of the first c. 1000 bp of the 28S rRNA (domain 1 of the 28S secondary structure) as well as COI. Cytochrome c oxidase subunit I amplification was done at the WAM using a BIO-RAD T100TM thermal cycler. Unpurified PCR products were sent to the Australian Genome Research Facility (Perth Node) for sequencing. Polymerase chain reactions for 28S were done at the Tick Pesticide Resistance Biotechnology Laboratory at the University of the Free State using a Corbett Research Gradient Palm-CyclerTM, with unpurified PCR products sent to Inqaba Biotechnical Industries in Pretoria for sequencing. Amplification reactions of 25 μl contained 2.5 μl of template DNA, 0.5μl of both the forward and reverse primers at 1μM each, and 21.5μl TopTaq Master Mix (Qiagen, R.S.A.).
The PCR process involved the following:
| Step | Process | Temperature | Time | Repeats
|
| 1 | Denaturation | 95oC | 5 min | 1x |
| 2 | Denaturation | 95oC | 30 s |
34x |
| 3 | Annealing | 45oC (28S), 46.2oC (COI) | 30 s | |
| 4 | Extension | 72oC | 1 min | |
| 5 | Final Extension | 72oC | 10 min | 1x |
| 6 | Hold | 20oC | ∞ | 1x |
Double-stranded PCR products were verified via agarose gel electrophoresis (1% agarose) and sent unpurified for sequencing.
Sequence editing and phylogenetic analysis
Cromatograms in .abi format were edited in Geneious R6 version 6.0.4 (http://www.geneious.com/). Sequence terminals were trimmed of low quality bases and overlapping segments were assembled. Before alignment, BLAST searches were done via the NCBI website (http://ncbi.nlm.nih.gov/) to check for putative contamination.
The same software was used to perform Geneious, MUSCLE and ClustalW alignments, as well as concatenation for the combined phylogenetic tree. For outgroups, sequences for Afrochthonius godfreyi (Ellingsen, 1912) (Pseudotyrannochthoniidae), Anagarypus heatwolei Muchmore, 1982 (Garypidae) and Synsphyronus apimelus Harvey, 1987 (Garypidae) were obtained from GenBank. Afrochthonius godfreyi acted as the root taxon, while the two garypid species served as the sister group to the Geogarypidae to assess whether or not the group is monophyletic. Alignments were produced for COI, 28S as well as the concatenated COI-28S tree.
Tests for maximum likelihood, neighbour-joining, minimum evolution, and maximum parsimony were performed using Mega 5 version 5.2.2 (http://www.megasoftware.net/). Mega 5 was chosen due its ease of use and incorporation of multiple phylogenetic tests, thus eliminating the need for multiple programs and the use of multiple file formats (Hall 2013). All tests were done with 1000 Bootstrap replications. A final Bayesian analysis was also run on the concatenated COI-28S tree using the MrBayes version 3.2.5 software package and the resulting consensus tree was viewed and edited in Fig Tree version 1.4.2. Trees were then edited in Adobe Photoshop CS5 to italicise species names, before being saved as .png files.
Systematics of the Geogarypidae
Family Geogarypidae Chamberlin, 1930
Garypinae Simon, 1879: 42; Tömösváry, 1882: 208; Balzan, 1892: 534.
Garypidae Simon: Hansen, 1893: 231; Ellingsen, 1904: 2; With, 1906: 89.
Geogarypinae: Chamberlin, 1930: 609; Beier, 1932: 227; Murthy & Ananthakrishan, 1977:
- [Type genus Geogarypus Chamberlin, 1930].
Geogarypidae Chamberlin: Harvey, 1986: 754; Harvey, 1992: 1420.
Diagnosis
According to Harvey (1992), the Geogarypidae possess the following apomorphies: pit-like structures present on the exterior margin of fixed chelal finger and the presence of a simple, single-bladed rallum without spinules. Further diagnostic characters include the possession of a sub-triangular carapace lacking any alae; two pairs of eyes situated on ocular tubercles located roughly one-third the length of the carapace from its anterior margin; the presence of a venom apparatus on both chelal fingers; and most species, with the exception of Geogarypus connatus Harvey, 1986 and A. castigatus sp. nov., possessing diplotarsate adults. The Geogarypidae can furthermore be separated from the family in which they were previously placed, the Garypidae, by the following: the position of trichobothrium isb; the presence of a spiracular stigmatic helix; a cheliceral rallum composed of a single blade lacking any spinules; coxa IV roughly as wide as coxa I; possessing a subterminal anal plate without a lateral rim that is not distinctly oval (Harvey 1986); and sternite XII without setae (this study, see below).
Description
Geogarypidae colouration varies from uniformly medium brown, as in A. triangularis comb. nov., to brown with light cream patches of various sizes on both the carapace (e.g. Geogarypus deceptor sp. nov.) and abdominal tergites (e.g. G. flavus stat. nov.). All South African geogarypids possess darkened spots medially on abdominal tergites I and II, as well as paired spots on tergites IV to X. Tergite III lacks a dark spot, but may present cream patches.
Chelicerae have five acuminate setae on the cheliceral palm (Fig. 1), with seta es originating at the base of the movable finger, bs located on a ridge centrally on the dorsal surface, setae sbs, ls and is grouping at the base of the fixed cheliceral finger, and gs located just above the base of the galea, on the movable finger. Lamina exterior and serrula interior present on fixed chelal finger. Serrula exterior present on the movable finger, consisting of multiple lamellae; two lamellae closest to the rallum sickle-shaped (Fig. 2). The rallum is comprised of a simple, single blade without any spinules. Galea either complex, with multiple rami curving ventrally (♀) (Fig. 3), or simple, consisting of a single spinneret usually without spinules (♂) (Fig. 4).
Carapace strongly sub-triangular in shape, constricting into a cucullus anteriorly, granulate in texture, and with a narrow furrow located posterior to the eyes (Fig. 5). Two pairs of corneate eyes situated on ocular tubercles located roughly one-third the length of the carapace from its anterior margin (Fig. 6). Pedipalpal coxae with distinct shoulder, trochanter with ventral apophysis. Femora and patellae granulate in texture, without any sensory trichobothria. Lyrifissures situated dorsally, usually on a raised surface, near base of the patellae. Surface of the chelal hand granulate, becoming smooth at base of chelal fingers. Venom apparatus and lamina defensor present on both chelal fingers. Fixed finger either with or without accessory teeth, usually with eight trichobothria, but A. castigatus sp. nov. with seven. Movable finger with four trichobothria. All trichobothria acuminate, with usual areolate shape. Fixed finger possessing a row of pit-like structures with raised rims on the exterior dorsal surface, each containing a central pore (Figs 7 & 8).
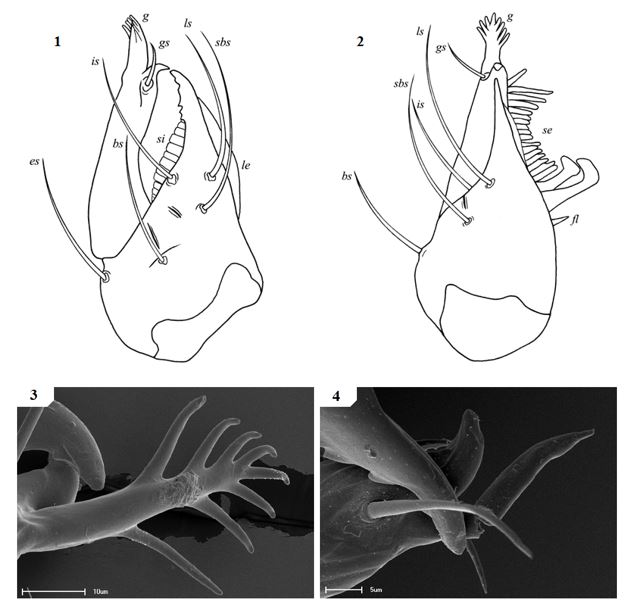
Figures 1–4. General morphology of geogarypid chelicerae galea. 1. Left chelicerae (female) dorsal view; 2. Same, prolateral view; 3. Geogarypus octoramosus sp. nov., female galea; 4. Same, male galea. For abbreviations of chela trichobothria see text.
Coxae of legs IV not significantly wider than coxae of legs I. Legs heterofemorate, with all joints of leg I and II movable. Legs I and II usually diplotarsate, though those of A. castigatus sp. nov. monotarsate, with arolium longer than claws. Abdomen sub-ovate in shape and broader than carapace. Tergites and sternites usually not divided, but a faint suture may be present ventrally, especially near the genital opercula. Pleural membrane wrinkled-plicate, with numerous investing setae present, set in small cuticular plates. In A. triangularis comb. nov. and A. castigatus sp. nov. males the cuticular plates are enlarged to form lateral sclerites. Spiracles open laterally of sternites III and IV, each with associated setae and a stigmatic helix (Harvey 1986).

Figures 5–8. Scanning electron micrographs of Afrogarpus excelsus (Beier, 1964) stat. nov. morphology. 5. Female carapace dorsal view; 6. Female ocular tubercle; 7. Male pit structure on retrolateral surface of fixed chela finger with pore; 8. Male fixed chelal finger showing pit structures on retrolateral surface.
Female genital opening not visible, sternite III not curved, with few setae (Fig. 9). Genital opercula of males with visible opening, sternite III curving posteriorly around opening, usually with many associated setae (Fig. 10). Anal cone situated sub-terminally, formed by fusion of tergite XII and sternite XII. Tergite XII bearing two associated setae, sternite XII with none.
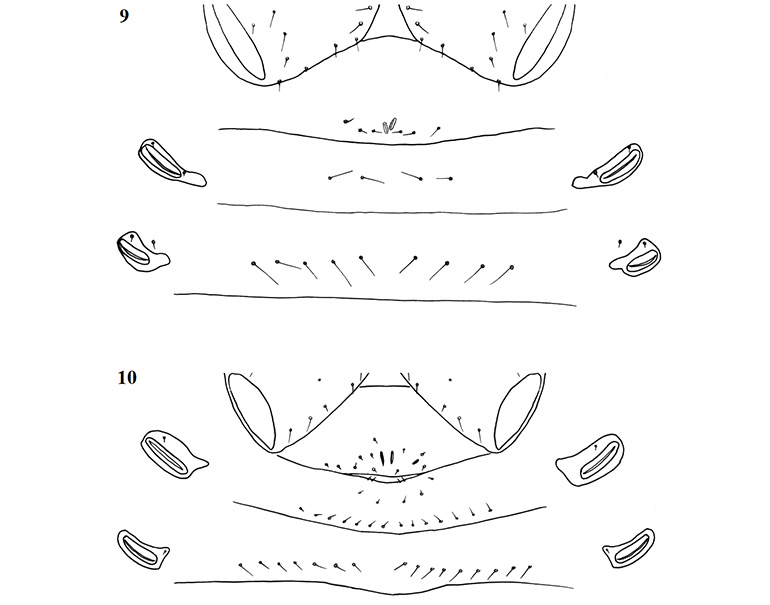
Figures 9–10. General morphology of geogarypid genital opercula. 9. Female genital area with spiracles; 10. Same, male.
Genus Afrogarypus Beier, 1931
Afrogarypus Beier, 1931: 317; Harvey, 1986: 758; Harvey, 1992: 1420
[Type species: Garypus senegalensis Balzan, 1892].
Geogarypus (Afrogarypus) Beier: Beier, 1932: 236; Beier, 1947: 320; Beier, 1955: 301.
Diagnosis
Originally distinguished by the presence of a deep dorsal sulcus on the chelal hand and lack of accessory teeth (Beier 1931; Harvey 1986); data generated in this study necessitates an update of the diagnosis of the genus. The following alterations are proposed:
i – Phylogenetic analysis indicated that A. minutus comb. nov., A. purcelli comb. nov., A. robustus comb. nov. and A. triangularis comb. nov. (until recently all in Geogarypus) group within the Afrogarypus clade. Morphological analysis of the above species showed the presence of a shallow depression on the dorsal surface of the chelal hand, a character not present in any of the species in the Geogarypus clade.
ii – Morphological analysis of species within the South African Afrogarypus clade indicated that the genus possesses accessory teeth. Some of the examined Afrogarypus species, such as A. excelsus (Beier, 1964) stat. nov. and Geogarypus species, such as Geogarypus octoramosus sp. nov., also share a small accessory tooth just above the first teeth on the exterior dorsal surface of the fixed chelal finger (Figs 11 & 12). The presence of accessory teeth is thus not a synapomorphy of any of the genera, and it is proposed that the presence of accessory teeth represents the pleisiomorphic state.
Species of the genus Afrogarypus can thus be distinguished from other South African Geogarypidae by the presence of either a well developed sulcus (Figs 13 & 14), or a concave depression on the dorsal surface of the chelal hand (Figs 15 & 16), resulting in a dorsal bulge at the base of the fixed chelal finger. With the exception of Afrogarypus castigatus sp. nov., which possesses a reduced compliment of 7/4, all the South African members of the genus possess the full 8/4 complement of trichobothria. Accessory teeth are present in some species.
Discussion
South African Afrogarypus possess some of the most morphologically distinct chela of all the Geogarypidae, with the presence of a dorsal sulcus being the most distinct feature. Phylogenetic analysis and subsequent combinations into the genus suggests that all but two of the originally described South African Geogarypidae, namely G. olivaceus and G. flavus stat. nov., belong to this genus.
South African species included
Afrogarypus carmenae sp. nov.; A. castigatus sp. nov.; A. excelsus (Beier, 1964) stat. nov.; A. impressus (Tullgren, 1907); A. megamolaris sp. nov.; A. minutus (Tullgren, 1907) comb. nov.; A. purcelli (Ellingsen, 1912) comb. nov.; A. robustus (Beier, 1964) comb. nov.; A. subimpressus (Beier, 1955) and A. triangularis (Ellingsen, 1912) comb. nov.
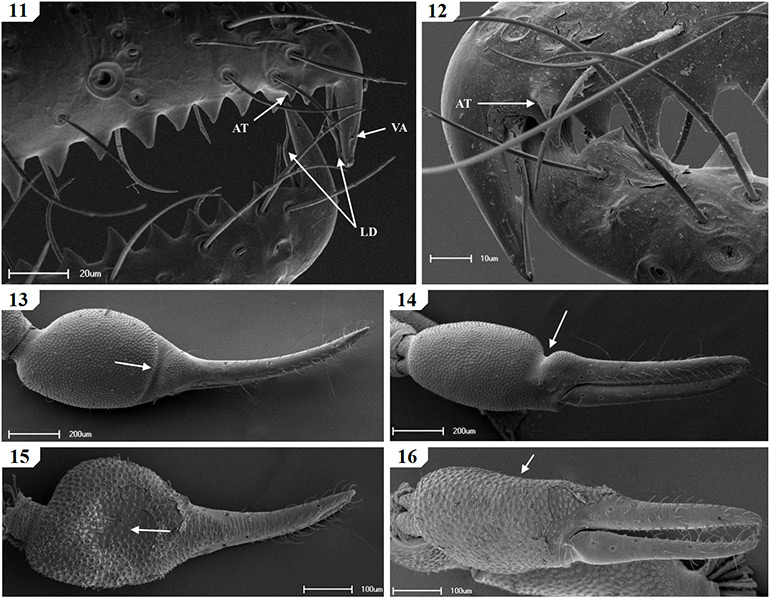
Figures 11–16. Scanning electron micrographs of South African Geogarypidae morphology. 11. Geogarypus octoramosus sp. nov., 12–14; Afrogarypus excelsus (Beier, 1964) stat. nov.; 15–16. Afrogarypus triangularis (Ellingsen, 1912) comb. nov.; 11–12. Exterior lateral view of the distal ends of the chela fingers, showing venom apparatus [VA], lamina defonsor [LD] and accessory tooth [AT]; 13, 15. Dorsal view of female right chela; 14, 16. Same, retrolateral view.
Geogarypus Chamberlin, 1930
Geogarypus Chamberlin, 1930: 609; Beier, 1932: 227; Beier, 1963: 241; Murphy &
Ananthakrishan, 1977: 104; Harvey, 1986: 760; Harvey, 1992: 1420
[Type species: Garypus minor Koch, 1873].
Diagnosis
With the revised diagnosis presented in this study, Geogarypus can easily be distinguished from members of Afrogarypus by the lack of any dorsal sulcus or concave depression on the chelal hand, instead presenting a continuous convex dorsal surface from the chelal stem to the base of the fixed finger (Figs 17 & 18). Accessory teeth may be present or absent.
Discussion
The only previously described South African species to remain in Geogarypus are G. olivaceus and G. flavus stat. nov. Pedipalps are surprisingly uniform in shape, with the most distinct differences found between the morphology of the chelal teeth of females. Males of this genus can be troublesome to distinguish due to the many variations in both size and colouration of a single species. As such, characters used within the proceeding identification key rely mainly on characters found in females. An example of the above can be seen when comparing the teeth at the base of the chelal fingers of both females (Figs 19 & 21) and males (Figs 20 & 22) of G. olivaceus and G. flavus stat. nov.

Figures 17–18. Scanning electron micrographs the female right chela of Geogarypus octoramosus sp. nov. 17. Dorsal view; 18. Retrolateral view.
South African species included
Geogarypus deceptor sp. nov.; G. flavus (Beier, 1947) stat. nov.; G. liomendontus sp. nov.; G. modjadji sp. nov.; G. octoramosus sp. nov.; G. olivaceus (Tullgren, 1907); G. tectomaculatus sp. nov.; G. variaspinosus sp. nov.
Key to the Geogarypidae genera and species of South Africa
1 Fixed chelal finger with trichobothria isb; legs I and II diplotarsate……..………………….…..2
1’ Fixed chelal finger without trichobothria isb; legs I and II monotarsate.
(Afrogarypus castigatus sp. nov.)…………………………………………………..………..p xx
2(1) Dorsal surface of chela hand with a well developed sulcus or concave region………….……..3
2’ Dorsal surface of chela hand convex in shape from stem to base of fixed finger. (Geogarypus)………………….………………………………………………………………11
3(2) Dorsal surface of chela hand with a well developed sulcus.
(Afrogarypus in part)……….………………………….………………………………………..4
3’ Dorsal surface of chela hand with a concave region, but not a distinct sulcus………….……..7
4(3) Dorsal sulcus narrow and deep; dorsal surface of chela before sulcus elevated above bulge at the base of the fixed finger………………………………………………………………………………5
4’ Dorsal sulcus wide and more shallow; dorsal surface of chela before sulcus on the same level as the bulge at the base of the fixed finger…………………………………………….………………..6
5(4) Chelal fingers distinctly longer than hand with stem.
(Afrogarypus excelsus stat. nov.)……………………………………………………..…….p xx
5’ Chelal fingers shorter than hand with stem.
(Afrogarypus impressus)…………………………………………………………..……….p xx
6(4’) Interior surface of chelal hand strongly covex; female galea with eight rami.
(Afrogarypus subimpressus)……………………….…………………………………..…..p xx
6’ Interior surface of chelal hand almost straight, giving the chela a chisel shape; female galea with nine rami.
(Afrogarypus megamolaris sp. nov.)……………………….…………………………..….p xx
7(3’) Chelal fingers distinctly longer than hand with stem.
(Afrogarypus purcelli comb. nov.)……..………………………………………………..….p xx
7’ Chelal fingers as long as hand with stem or shorter…………………………………….………8
8(7’) Chelal fingers more or less as long as hand with stem…………………………………….…….9
8’ Chelal fingers distinctly shorter than hand with stem……………………………………….…10
9(8) First tooth behind venom apparatus noticeably more sclerotised and slightly larger than the teeth just after it; female galea with nine rami.
(Afrogarypus carmenae sp. nov.)…………………………………………………….….p xx
9’ Chelal teeth not as above; female galea elongate and with eight rami.
(Afrogarypus minutus comb. nov.)……………………………………………………..…p xx
10(8’) Chela long and slender; trichobothrial distribution normal, not grouping together.
(Afrogarypus robustus comb. nov.)………………………………………….…….…..……p xx
10’ Interior surface of chelal hand strongly convex, giving the chela a triangular appearance; trichobothria eb, esb, est, ib, isb, b, sb and st grouping proximally within the first third of the finger length, separated roughly by another third of the finger length from the distal group, consisting of trichobothria ist, it, et and t.
(Afrogarypus triangularis comb. nov.)………………………………………………………p xx
11(2’) Female galea with nine rami……………………………………………………………..…….12
11’ Female galea with fewer than nine rami………………………………………………..………17
12(11) Movable chelal finger with a distinct lone tooth at its base.
(Geogarypus deceptor sp. nov.)……………………………………………………………….p xx
12’ Movable chelal finger without a basal tooth separated from rest of teeth…………………………..13
13(12’) All teeth separate with no fusion………………………………………………………………………………….14
13’ Some teeth fused at the base of the chelal fingers…………………………………………………………..16
14(13) Male and female with ≥40 teeth on the fixed chelal finger; first three basal teeth on movable finger separated by one tooth distance from each other; male galea with one or more spinules.
(Geogarypus flavus stat. nov.)…………………………………………………………………………………p xx
14’ Male and female with <38 teeth on the fixed chelal finger; first three basal teeth on female movable finger adjacent to each other, without any gaps…………………………………………………15
15(14’) First three basal teeth on female’s movable finger grouped very close to each other, almost fused; said basal teeth originating above trichobothria b in females.
(Geogarypus olivaceus)………………………………………………………………………………………….p xx
15’ First three basal teeth on female’s movable finger grouped close at the base of the teeth, but teeth distinctly separate; said basal teeth originating between trichobothria b and sb in females.
(Geogarypus tectomaculatus sp. nov.)…………………………………………………………………….p xx
16(13’) First four basal teeth on both female chelal fingers fused.
(Geogarypus liomendontus sp. nov.)………………………………………………………………………..p xx
16’ Only first three basal teeth fused on fixed chelal finger of female; no fused teeth on movable finger, though first three basal teeth grouped close together.
(Geogarypus modjadji sp. nov.)……………………………………………………………………………..p xx
17(11’) Female galea with eight rami.
(Geogarypus octoramosus sp. nov.)………………………………………………………………………..p xx
17’ Female galea with variable number of rami, ranging from five to seven.
(Geogarypus variaspinosus sp. nov.)………………………………………………………………………p xx
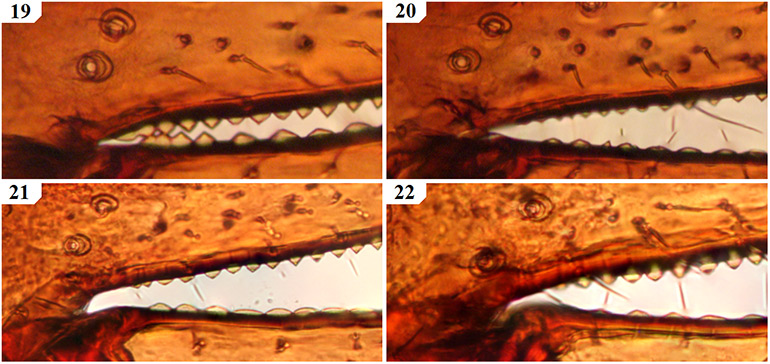
Figures 19–22. Tooth morphology at the base of the chelal fingers of Geogarypus spp. showing distinct differences in females, with males mainly differing with regard to teeth size between species. 19. Female Geogarypus olivaceus (Tullgren, 1907); 20. Same, male; 21. Female Geogarypus flavus (Beier, 1947) stat. nov.; 22. Same, male.
Afrogarypus carmenae sp. nov.
Figs 23-35
Holotype: ♀, SOUTH AFRICA, Western Cape, Clanwilliam, Gecko Creek Wildlife Lodge, 32o 23’S, 18o 59’E, 331 m a.s.l., Bushveld, Leaf litter sifting, leg. J.A. Neethling, 21.XII.2012 (NMBA P00228).
Paratypes: 1♀, 3♂ (NMBA P00229), Same data as holotype.
Etymology
Named after Carmen Neethling (née Luwes) who, throughout the study, helped to collect many of the newly described species.
Diagnosis
Medium sized species with both males and females reaching a total length (cucullus to posterior abdominal margin) of 2.14 mm. Carapace uniform brown to light brown in both sexes. All pedipalp segments similar in colouration to the carapace. Concave depression present on dorsal surface of chela. Abdominal tergites mostly medium brown with dark patches, though males can possess cream coloured markings. Abdominal sternites light brown in both sexes, weakly sclerotised in females and strongly sclerotised throughout in males. Pedipalp coxae brown in colour with distinct shoulder, legs I-IV as well as remaining coxae tan to pale yellow. Though mostly equal in length, females tend to be more bulky with larger abdomens, while males are slimmer and more elongate.
Specimens of A. carmenae sp. nov. resemble A. purcelli comb. nov. in general appearance, but can be distinguished by lacking the inclusion on the prolateral surface of the chela present in A. purcelli comb. nov.
Description
Carapace: Strongly sub-triangular, narrow furrow posterior to the eyes (Fig. 33). Overall brown to light brown in both sexes, medial furrow and posterior margin somewhat lighter. Uniformly granular throughout, heavily constricted anteriorly into cucullus, constriction beginning at medial furrow. Two pairs of corneate eyes situated on ocular tubercles, located about one-third away from anterior edge. Four prominent setae located on anterior edge, row of 10 setae, seated within rims, located on posterior margin. Numerous small setae present on the carapace.
Abdomen: Abdominal tergites brown to light brown with dark and cream patches in both sexes, granular. Tergites I and II with large median dark spot each. Tergite III without dark patches. Tergites IV-X with paired indistinct dark patches, just lateral of the midline. Light coloured specimens present indistinct, vague, spots. Tergites XI and XII without patches. Sternites light brown with indistinct dark patches in both sexes, weakly sclerotised in females, well sclerotised throughout in males. Pleural membrane wrinkled-plicate, cream.
Pedipalp: Trochanter rounded, slightly elongate in shape, granular, same colour as carapace. Small setae scattered over entire surface, except stem. Ventral apophysis present. Pedipalp femur (Fig. 28) same colour, granular. Narrow at base, widening evenly before it is constricted at distal end. Pedipalp patella (Fig. 28) same colour as previous, granular. Narrow, slightly angled at base, widening evenly into elongate cone. Lyriform fissures visible just anterior of base, located on a bulge, dorso-prolaterally on patella.
Chela: (Figs 23-28) Uniformly brown, may be slightly darker than rest of pedipalps. Granular from stem to base of fingers. Both fingers smooth, slightly shorter than hand (with pedicel). Concave depression present on dorsal surface of chela, resulting in small bulge at base of fixed finger. Chela hand weakly convex on the prolateral edge, retrolateral edge mostly straight.

Figures 23–26. Digital microscope photographs of Afrogarypus carmenae sp. nov. right chela: Female (23, 24) and Male (25, 26). 23, 25. Retrolateral view; 24, 26. Dorsal view. Scale bar: 1.00 mm.
Fixed chela finger with eight trichobothria, movable chela finger with four, both fingers narrow, curving slightly to interior. Venom apparatus present on both fingers. Trichobothria eb and esb situated on retrolateral surface of the fixed chelal finger, located just above base of finger, one trichobothrial width between them. Trichobothria ib located on prolateral surface, anterior and dorsal of esb, est located opposite and anterior, with one trichobothrial width, of ib. Trichobothria isb located roughly halfway along fixed finger, situated dorsally, ist is located on prolateral surface, approximately one third from anterior edge of fixed chelal finger. Trichobothria it located dorsally, roughly one fifth from anterior edge of fixed chelal finger, et located on retrolateral surface, just distal of it. Regarding movable finger, trichobothria b and sb located on retrolateral surface, just forward of finger base. Trichobothria st located on same surface, situated between est and isb, t located between ist and it. Chelal teeth acute, slightly retrorse. Fixed finger with 39 teeth (♀), 35 (♂). First tooth just before venom apparatus slightly enlarged, strongly sclerotised, followed by seven (♂) or eight (♀) smaller, closely spaced, roughly equal sized teeth before the first large accessory tooth (Fig. 32). Further teeth slightly retrorse and acute, arranged roughly in two rows. Movable finger with 31 teeth (♀), 29 (♂), tooth just before venom apparatus being very small. Following eight (♀) or 10 (♂) teeth after small tooth acute, retrorse, further teeth reduced into small teeth plates. In females, last five teeth increase in size into distinct group, spaced tightly against each other. In males, rear teeth may increase in size slightly, or remain as acute points.
Chelicera: Hand with five setae as in Fig. 1. Fixed cheliceral finger with six teeth arranged as in Fig. 29. Galea complex, with nine rami in a fan shape as in Fig. 30 (♀) or simple, no rami (♂) (Fig. 31). Rallum composed of a single blade in both sexes. Serrula exterior with 19 lamella (♀) or 17 (♂). Lamina exterior present in both sexes.
Coxae and legs: Pedipalp coxae light brown, shoulder distinct. Legs I-IV, including coxae, tan to pale yellow. All legs diplotarsate (Figs 34 & 35).
Measurements (mm): body length ♀ 2.14 ♂ 2.14; carapace ♀ 0.74 x 0.69 (1.07) ♂ 0.61 x 0.51 (1.19). Palp femur ♀ 0.74 x 0.20 (3.70) ♂ 0.62 x 0.16 (3.87), patella ♀ 0.62 x 0.20 (3.10) ♂ 0.54 x 0.17 (3.17), hand ♀ 0.57 x 0.33 ♂ 0.47 x 0.25, chela ♀ 1.16 (3.51) ♂ 0.96 (3.84), movable finger ♀ 0.55 ♂ 0.49. Leg I femur ♀ 0.32 x 0.11 ♂ 0.26 x 0.10, patella ♀ 0.19 x 0.10 ♂ 0.15 x 0.09, tibia ♀ 0.26 x 0.07 ♂ 0.21 x 0.06, metatarsus ♀ 0.17 x 0.06 ♂ 0.15 x 0.05, tarsus ♀ 0.16 x 0.04 ♂ 0.15 x 0.04. Leg IV femur ♀ 0.15 x 0.12 ♂ 0.13 x 0.10, patella ♀ 0.50 x 0.20 ♂ 0.42 x 0.17, tibia ♀ 0.43 x 0.10 ♂ 0.35 x 0.09, metatarsus ♀ 0.22 x 0.07 ♂ 0.18 x 0.06, tarsus ♀ 0.19 x 0.05 ♂ 0.18 x 0.04.
Remarks
Many museum exemplars were found to be misidentified as either immature A. robustus comb. nov. or A. purcelli comb. nov. The type specimens were sampled together with a group of 13 individuals congregating under shaded vegetation along a rocky ridge. No immature specimens were sampled. The species has been collected in Fynbos and Nama Karoo biomes.
Additional material examined
SOUTH AFRICA: Western Cape: 1♀, 2♂, Bredasdorp, De Hoop Nature Reserve, 34o 28’S, 20o 30’E, 26 m a.s.l., Coastal Fynbos, Leaf litter sifting, leg. J.A. Neethling & C. Luwes, 24.XII.2013 (NMBA P00227); 1♀, Bredasdorp, De Hoop Nature Reserve, Potberg, 34o 22’S, 20o 32’E, 180 m a.s.l., leg. C.R. Haddad, 7.IV.2004 (WAM T79013); 1♀, Cape Town, Table Mountain, 33o 58’S, 18o 23’E, 15.I.2007 (WAM T85959); 1♀, Clanwilliam, Van Rhynsdorp Road [32o 10’S, 18o 2’E], leg. Leipoldt, XII.1997 (SAMC 9112); 1♀, 1♂, Hermanus, Fernkloof Nature Reserve,
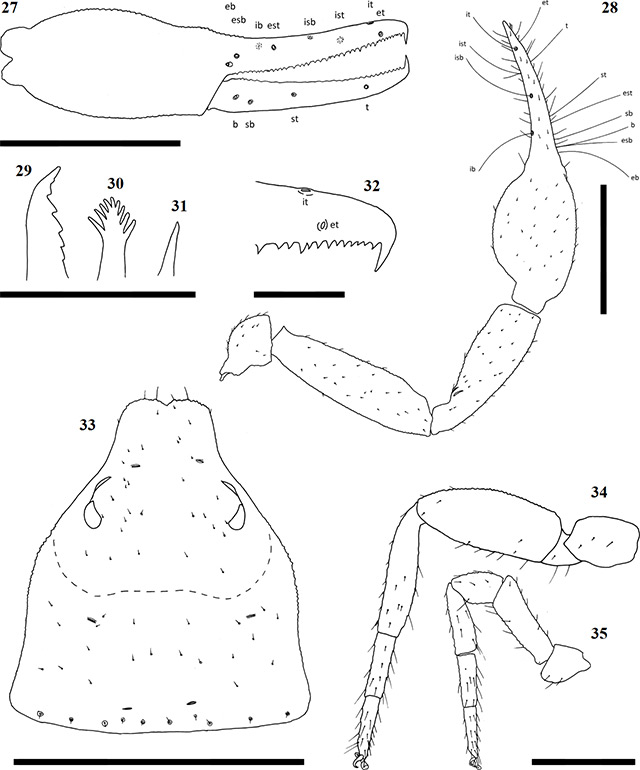
Figures 27–35. Afrogarypus carmenae sp. nov. 27–29, 31–35. Female; 30. Male; 27. Right chela, retrolateral view; 28. Right pedipalp, dorsal view; 29. Chelicera movable finger, dorsal view; 30, 31. Galea; 32. Right chela fixed finger, anterior teeth arrangement; 33. Carapace, dorsal view; 34. Right leg IV, prolateral view; 35. Right leg I, prolateral view. Scale bars: Figs 27, 28, 33-35: 0.50 mm; Figs 29-32: 0.10 mm.
34o 24’S, 19o 16’E, leg. R. Lyle, 29.IX.2007 (WAM T81146, T86777); 1♀, Kleinmond [34o 20’S, 19o 01’E], leg. H.G. Robertson, 4.XI.1995 (SAMC ENW-COO 6599); 1♀, 1♂ Laingsburg, Anysberg Nature Reserve [33o 28’S, 20o 35’E], leg. C.R. Haddad, 24.IX.2007 (WAM T86782, T86784); 1♀, Pringle Bay, Cape Hanglip [34o 23’S, 18o 50’E], leg. R.F. Lawrence, XII.1962 (NMSA 7897); 1♀, 1♂, Simon’s Town, Cape of Good Hope Nature Reserve, Smitswinkelvlakte [34o 17’S, 18o 26’E], leg. H.G. Robertson, X.1998 (SAMC ENW-COO 5405, 5406); 2♀, Simon’s Town, Cape of Good Hope Nature Reserve, Teeberg [34o 14’S, 18o 24’E], leg. H.G. Robertson, X.1998 (SAMC ENW-COO 5409, 5435); 1♀, Stellenbosch [33o 55’S, 18o 52’E], leg. J. Botha, 19.V.1990 (NCA 90.286).
Afrogarypus castigatus sp. nov.
Figs 36-51
Holotype: ♀, SOUTH AFRICA, KwaZulu-Natal, St. Lucia, St. Lucia Coastal Forest, 28o 23’S, 32o 24’E, 20 m a.s.l., Coastal forest, Canopy fogging Trichilia emetica (Natal Mahogany), leg. J.A. Neethling & C. Luwes, 13.V.2012 (NMBA P00219).
Paratypes: 1♀, 3♂, SOUTH AFRICA, KwaZulu-Natal, St. Lucia, iSimangaliso Wetland Park, Crocodile Centre, 28o 21’S, 32o 25’E, 24 m a.s.l., Indigenous forest patch, Canopy fogging Breonadia salicina (Matumi), leg. J.A. Neethling & C. Luwes, 14.V.2012 (NMBA P00220).
Etymology
Castigatus (Latin adj. meaning compact). The species is named for its unusual morphological features, including reduced trichobothrial count on the fixed chelal finger and monotarsate legs I and II.
Diagnosis
Small species. Females reach lengths (cucullus to posterior abdominal margin) of 1.55 mm, while males reach 1.35 mm in length. Carapace dark brown in both sexes, with the medial furrow and posterior light brown to cream in colour. All pedipalp segments follow the same colouration as the carapace. Concave depression present on dorsal surface of chela. Abdominal tergites light brown to cream with dark patches in both sexes. Abdominal sternites light brown to cream in both sexes, with no markings; weakly sclerotised in females and partly to well sclerotised in males. Legs I and II monotarsate and cream to light brown in both sexes, with corresponding coxae cream in colour.
Currently the smallest geogarypid species in South Africa, with very little sexual dimorphism in morphology and colouration, though males tend to be smaller than females. The species resembles small A. triangularis comb. nov. and A. subimpressus in overall appearance, but can be easily distinguished by having a reduced trichobothrial compliment of 7/4, due to the absence of trichobothria isb, as well as having monotarsate leg pairs I and II. The species resembles A. triangularis comb. nov. in particular, due to the presence of a very convex prolateral surface on the chela, a concave dorsal chelal depression and the presence of pleural sclerites on the abdomen of adult males.
Description
Carapace: Strongly sub-triangular, broader than long in both sexes, with narrow furrow posterior to the eyes (Fig. 48). Generally dark brown in colour, medial furrow and posterior of carapace light brown to cream in both sexes. Uniformly granular, heavily constricted anteriorly into cucullus, constriction beginning at the medial furrow. Two pairs of corneate eyes situated on ocular tubercles, located ca. one third away from anterior edge. Four prominent setae located on anterior edge, row of nine setae located on posterior margin. Numerous small setae present on carapace.
Abdomen: Very compact and almost circular in appearance. Abdominal tergites granular, light brown to cream, with darker patches in both sexes. Tergites I and II with large median dark spot each, flanked by light coloured patches, in turn flanked by light brown patches at lateral edges. Tergite III without dark medial patch, cream, flanked at lateral edges by light brown markings. Tergites IV-X with paired, indistinct dark patches just lateral of midline, flanked at lateral edges by light brown markings. Tergites XI and XII without dark patches. Males possess lateral sclerites on pleural membrane (Fig. 49) as follows; one each below tergites III and IV, two below tergite V, four sclerites in two rows of two below tergite VI, four sclerites in single row below tergite VII, five sclerites in two rows of three and two below tergite VIII, three in single row below tergite IX, one below tergite X. Sternites light brown to cream in both sexes, no markings, weakly sclerotised in females, partly to well sclerotised in males. Pleural membrane wrinkled-plicate, cream, with light brown lateral sclerites, in males.
Pedipalp: Trochanter rounded in shape, granular, coloured same as carapace. Small setae scattered over entire surface, with the exception of the stem. Apophysis located ventrally. Pedipalp femur (Fig. 43) same colour and granular. Narrow at stem, widening quickly to form base, then widening slightly before constricting again at end. Pedipalp patella (Fig. 43) same colour as femur, granular. Narrow, very angled at base, widening evenly into short cone. Lyriform fissures visible just anterior of base, located on bulge, dorso-prolaterally on patella.
Chela: (Figs 36–39, 42, 43) Same colour as carapace, uniformly granular from stem to base of the movable fingers, granular over first third of dorsal surface of fixed finger. Both fingers much shorter than hand (with pedicel). First third of fixed chelal finger with characteristically swollen dorsal area. Concave depression present on dorsal surface of chela, resulting in small bulge at base of fixed finger. Chela hand strongly convex on prolateral edge, retrolateral edge slightly convex.

Figures 36–39. Digital microscope photographs of Afrogarypus castigatus sp. nov. right chela: Female (36, 37) and Male (38, 39). 36, 38. Retrolateral view; 37, 39. Dorsal view. Scale bar: 1.00 mm.
Fixed chela finger with seven trichobothria (isb absent), movable chela finger with four, both fingers narrow, curving slightly to the interior. Venom apparatus present on both fingers. Trichobothria eb and esb situated on retrolateral surface of fixed chelal finger, just above base of finger, one trichobothrial width between them. Trichobothria ib on prolateral surface, distal of esb by just more than one trichobothrial width; est opposite, directly anterior of ib. Trichobothria isb absent, ist on prolateral surface, approximately half way from anterior edge of fixed chelal finger, situated close to chelal teeth. Trichobothria it located dorsally, just over one-quarter finger length from anterior edge of fixed chelal finger, et on retrolateral surface, just distal of it. On the movable finger, trichobothria b, sb and st group together with equal distances from each other below group formed by eb, esb and est. Trichobothria t located roughly one-third finger length from anterior edge of movable finger. Chelal teeth acute and retrorse. Fixed finger with 19 teeth in both sexes. First three teeth just before the venom apparatus slightly fused and raised (Fig. 44), remaining teeth spaced widely apart, decreasing in size to base of fixed finger. Movable finger with 16 teeth (♀), 15 (♂), two teeth just before venom apparatus being small and raised. Remaining teeth retrorse, facing backward, decreasing in size to base of moving finger.
Chelicera: Hand with five setae as in Fig. 1. Fixed cheliceral finger with four teeth arranged as in Fig. 45. Galea complex with nine rami as in Fig. 46 (♀), or simple with no rami (♂) (Fig. 47). Rallum composed of single blade in both sexes. Serrula exterior with 15 lamella in both sexes. Lamina exterior present in both sexes.
Coxae and legs: Pedipalp coxae same colour as carapace, with distinct shoulder. Legs I and II monotarsate (Figs 40 & 51), cream to light brown in both sexes, corresponding coxae cream. Legs III and IV diplotarsate (Figs 41 & 51), with same colouration as front legs.
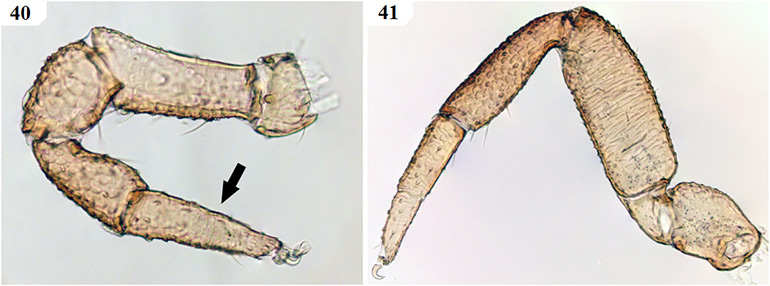
Figures 40–41. Digital microscope photographs of Afrogarypus castigatus sp. nov. female leg morphology. 40. Monotarsate leg I with arrow indicating the fusion of last two leg segments; 41. Diplotarsate leg IV.
Measurements (mm): body length ♀ 1.55 ♂ 1.35; carapace ♀ 0.55 x 0.62 (0.88) ♂ 0.50 x 0.57 (0.87). Palp femur ♀ 0.49 x 0.15 (3.26) ♂ 0.45 x 0.14 (3.21), patella ♀ 0.36 x 0.14 (2.57) ♂ 0.32 x 0.14 (2.28), hand ♀ 0.45 x 0.27 ♂ 0.41 x 0.26, chela ♀ 0.75 (2.77) ♂ 0.68 (2.61), movable finger ♀ 0.29 ♂ 0.26. Leg I femur ♀ 0.19 x 0.10 ♂ 0.18 x 0.09, patella ♀ 0.12 x 0.09 ♂ 0.12 x 0.08, tibia ♀ 0.16 x 0.07 ♂ 0.13 x 0.07, metatarsus + tarsus ♀ 0.20 x 0.05 ♂ 0.18 x 0.05. Leg IV femur ♀ 0.11 x 0.08 ♂ 0.09 x 0.07, patella ♀ 0.32 x 0.12 ♂ 0.30 x 0.11,
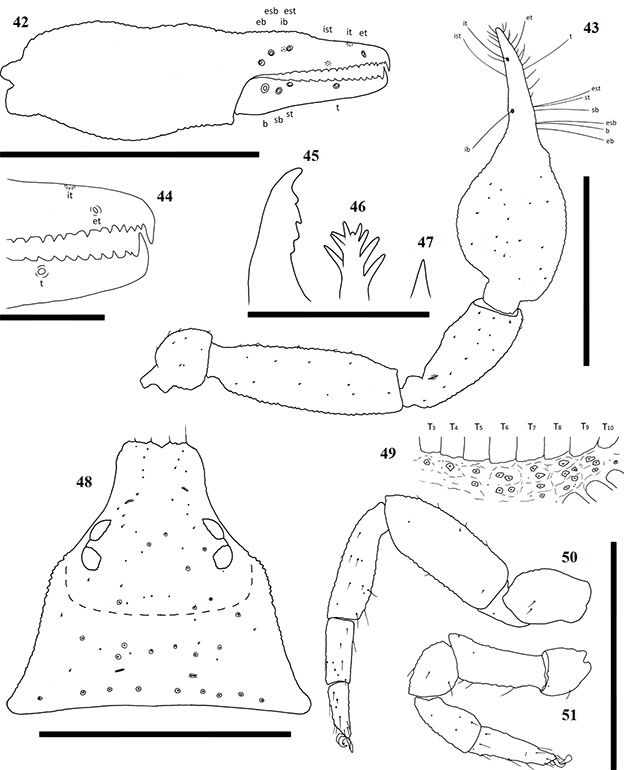
Figures 42–51. Afrogarypus castigatus sp. nov. 42–46, 48, 50, 51. Female; 47, 49. Male. 42. Right chela, retrolateral view; 43. Right pedipalp, dorsal view; 44. Anterior retrolateral view of chela fingers; 45, Distal end of chelicera movable finger, dorsal view; 46, 47. Galea; 48. Carapace, dorsal view. 49. Abdominal pleural sclerites; 50. Leg IV, prolateral view; 51. Leg I, prolateral view. Scale bars: Figs 42, 43, 48, 50, 51: 0.50 mm; Fig. 44: 0.10 mm; Figs 45-47. 0.05 mm.
tibia ♀ 0.27 x 0.08 ♂ 0.24 x 0.08, metatarsus ♀ 0.13 x 0.05 ♂ 0.12 x 0.05, tarsus ♀ 0.12 x 0.04 ♂ 0.11 x 0.04.
Remarks
Although this is the only South African geogarypid species with a reduced trichobothrial compliment, it is nonetheless not the only geogarypid displaying such a reduction. Geogarypus connatus from Australia, as well as G. bucculentus (Beier, 1955) from the Juan Fernandez Islands, Chile, also present the absence of trichobothria isb (Harvey 1987b). Adults of Afrogarypus castigatus sp. nov. and G. connatus also possess monotarsate legs I and II, while G. bucculentus adults are diplotarsate. Given their geographic locations, the three species may form a monophyletic group that originated in Gondwanaland (Harvey 1987b; Harvey 1996).
Additional material examined
SOUTH AFRICA: KwaZulu-Natal: 4♀, 6♂, St. Lucia, iSimangaliso Wetland Park, Meersig, 28o 14’S, 32o 29’E, 14 m a.s.l., Wetland, Canopy fogging Syzygium cordatum, leg. J.A. Neethling & C. Luwes, 14.V.2012 (morphological dissection, phylogenetic analysis and S.E.M.).
Afrogarypus excelsus (Beier, 1964) stat. nov.
Figs 52-64
Geogarypus excelsus Beier, 1964: 61, fig. 22 (Holotype ♀, South Africa, Eastern Cape,
Grahamstown [33o18’S, 26o31’E], leg. J. Hewitt, deposited in NMZA, not examined).
Geogarypus excelsus excellens Beier, 1964: 62, fig. 23 (2♀, 1♂, 2 Deutonymphs, Syntypes,
No collection information, NMZA, not examined; 4♀, 4♂, 1 Tritonymph, Paratypes, No collection information, NMZA, not examined).
Geogarypus (Afrogarypus) excelsus excellens (Beier, 1964): Beier, 1966: 461.
Afrogarypus excelsus excelsus (Beier, 1964): Harvey, 1986: 758; Harvey, 1991: 250;
Dippenaar-Schoeman & Harvey, 2000: 93 [stat. nov.].
Afrogarypus excelsus excellens (Beier, 1964): Harvey, 1986: 758; Harvey, 1991: 250;
Dippenaar-Schoeman & Harvey, 2000: 93 [syn. nov.].
Diagnosis
Large species. Females can reach a total length (cucullus to posterior abdominal margin) of 2.96 mm, with males reaching up to 2.27 mm. Carapace dark brown with light brown markings in the form of an hourglass located on the posterior margin and stretching up to the middle of the carapace. All pedipalp segments dark brown in colour. Abdominal tergites brown with dark patches, abdominal sternites light brown to tan and weakly sclerotised (♀), brown with dark patches and heavily sclerotized (♂). Pedipalp coxae medium brown in colour with distinct shoulder, legs I-IV as well as remaining coxae tan. As with many pseudoscorpions, the male of the species tends to vary in terms of size and colouration, while adult females tend to be similarly-sized within a population. Males are generally smaller and can be easily identified by their dark, sclerotised sternites and slender, elongate pedipalps.
Afrogarypus excelsus stat. nov. closely resembles A. impressus, but differs with regard to the chela ratio, finger length, as well as chelal teeth morphology. The most characteristic difference is that the chelal fingers of A. excelsus stat. nov. are longer than the hand-stem complex, while they are shorter than the complex in A. impressus. Together with A. impressus, A. excelsus stat. nov. is the only other large South African geogarypid to exhibit the presence of a well developed, deep dorsal sulcus. Afrogarypus subimpressus and A. megamolaris sp. nov. also possess a developed dorsal sulcus, but it is not nearly as well defined as those found in the former species. The latter two species further differ in size, as well as having chelal teeth that are spaced widely apart.
Description
Carapace: Strongly sub-triangular, narrow furrow posterior to the eyes (Fig. 60). Dark brown anterially, until beginning of the furrow, light brown marking in shape of an hourglass in posterior half. Uniformly granular, except for posterior margin. Heavily constricted anteriorly into cucullus, constriction beginning just anterior of the medial furrow. Two pairs of corneate eyes situated on ocular tubercles, located about one-third away from anterior edge. Four prominent setae located on anterior edge, row of eight setae located on posterior margin. Numerous small setae present on carapace, some situated within large rims similar to those of trichobothria.
Abdomen: Abdominal tergites brown (♀) or light brown (♂), with dark patches. Tergites I and II with large medially located dark spot each. Tergite III without dark patches. Tergites IV–X (♀) with paired dark patches, lateral of the midline, can be slightly lighter in colour (♂ may lack these paired patches). Tergites XI–XII without any patches. Sternites light brown to tan, weakly sclerotised (♀), brown with dark patches, heavily sclerotized (♂). Pleural membrane wrinkled-plicate with invested setae.
Pedipalp: Trochanter rounded, granular, uniformly dark brown with small setae located on the dorsal and dorso-lateral sides. Apophysis located ventrally. Pedipalp femur (Fig. 57) dark brown, granular. Narrow at base, widening evenly until just posterior of anterior margin, where it is constricted. Pedipalp patella (Fig. 57) dark brown, granular, narrow at base, widening quickly until anterior edge. Lyriform fissures visible just anterior of base, located on a bulge, dorso-prolaterally on patella.
Chela: (Figs 52–57) Uniformly brown, granular from stem to base of fingers. Both fingers smooth. Well developed dorsal sulcus present, located just posterior of the base of the fixed finger. Dorsal bulge located just anterior of sulcus. Fixed chela finger with eight trichobothria, movable chela finger with four. Both fingers long and narrow, curving to interior. Fingers just as long as, or slightly longer than, hand (with pedicel). Venom apparatus present on both fingers.
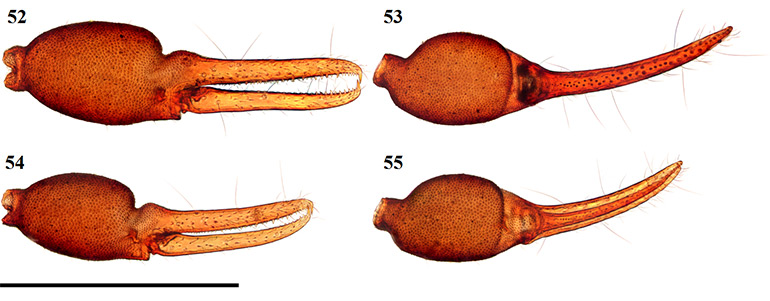
Figures 52–55. Digital microscope photographs of Afrogarypus excelsus (Beier, 1964) stat. nov. right chela: Female (52, 53) and Male (54, 55). 52, 54. Retrolateral view; 53, 55. Dorsal view. Scale bar: 1.00 mm.
Trichobothria eb and esb situated about one trichobothrial width from each other, located respectively retrolaterally and dorso-retrolaterally off base of fixed finger; ib situated about one-quarter anterior of finger base, located dorso-prolaterally; est opposite, just anterior of ib; isb situated almost dorsally, located about halfway along fixed finger, closer to est than to ist; ist located about one-quarter from the anterior edge of the fixed finger, situated prolaterally; it located dorsally, two widths along finger from ist; et located two widths from it, sits opposite on the same horizontal line with ist. Regarding the movable finger, b located just anterior of finger base, situated dorso-retrolaterally on finger in line vertically with eb; sb located about. two trichobothrial widths from b, situated slightly ventrally; st located about halfway along movable finger, closer to sb than t and situated slightly more dorsally; t located about one-quarter from anterior edge of movable finger, just forward of ist. Chelal teeth acute and retrorse, some curving back slightly. Fixed finger with 41 teeth (♀), 39 (♂). Six small, closely spaced teeth just behind venom apparatus, followed by large tooth (Fig. 61). Further teeth are arranged in two rows, reducing in size proximally, still acute. Movable finger with 26 teeth (♀), 25 (♂). Movable finger’s teeth nearly contiguous, reduced to small projections proximally, after first nine (♀) or seven teeth (♂).
Chelicera: Hand with five setae as in Fig. 1. Fixed cheliceral finger with five teeth arranged as in Fig. 62. Galea complex, with nine rami arranged evenly across the tip (♀) (Fig. 63), simple, with no rami (♂) (Fig. 64). Rallum composed of single blade in both sexes. Serrula exterior with 13 lamella (♀) or 17 (♂). Lamina exterior present in both sexes.
Coxae and legs: Pedipalp coxae brown, with distinct shoulder. Multiple long setae present along inner medial ridges. Coxae of legs I–IV, as well as all leg segments, tan. Trochanters of legs I and II compact and small, larger and elongate on legs III and IV. Femora of legs I and II longer than patellae and tibiae respectively (Fig. 58). Metatarsi and tarsi not fused, joint movable. Tarsal claws simple and curved, arolium much longer than claws, not divided. Femur-patella joint immovable in leg IV, tibia slightly curved, metatarsi and tarsi not fused and joint movable (Fig. 59). Claws and arolium similar to those of legs I–III.
Measurements: body length ♀ 2.96 ♂ 2.27; carapace ♀ 1.32 x 1.24 (1.06) ♂ 1.27 x 1.24 (1.02). Palp femur ♀ 1.34 x 0.39 (3.43) ♂ 1.40 x 0.41 (3.41), patella ♀ 1.07 x 0.33 (3.24) ♂ 1.07 x 0.35 (3.05), hand ♀ 0.75 x 0.39 ♂ 0.62 x 0.37, chela ♀ 1.52 (3.89) ♂ 1.31 (3.54), movable finger ♀ 0.75 ♂ 0.66. Leg I femur ♀ 0.39 x 0.16 ♂ 0.37 x 0.15, patella ♀ 0.24 x 0.15 ♂ 0.21 x 0.14, tibia ♀ 0.31 x 0.10 ♂ 0.28 x 0.09, metatarsus ♀ 0.20 x 0.08 ♂ 0.17 x 0.07, tarsus ♀ 0.15 x 0.05 ♂ 0.15 x 0.05. Leg IV femur ♀ 0.20 x 0.13 ♂ 0.21 x 0.14, patella ♀ 0.54 x 0.21 ♂ 0.57 x 0.23, tibia ♀ 0.43 x 0.13 ♂ 0.47 x 0.13, metatarsus ♀ 0.24 x 0.08 ♂ 0.25 x 0.08, tarsus ♀ 0.20 x 0.06 ♂ 0.20 x 0.06.
Remarks
The last of the South African geogarypid species to be described in the mid-20th century, A. excelsus stat. nov. was originally separated into two subspecies, namely A. excelsus excelsus and A. excelsus excellens. Beier (1964) based his decision on specimens with no collection information that he found together in the same series as the species’ holotype. These specimens differed from the holotype of A. excelsus as follows: slimmer and slightly more elongate chela, trichobothria ist on the fixed chela finger being slightly closer to it and thus forming a proximal group with et, and trichobothria st on the movable chela finger being slightly closer to sb than to t, whereas it is evenly spaced between the two in the holotype of A. excelsus. Specimens agreeing with both subspecies descriptions were collected together from multiple locations, including Fort Fordyce Nature Reserve and Pirie Forest Reserve. Phylogenetic analyses of these specimens clearly show that they are the same species, and thus the slight differences in morphology can be attributed to intraspecific variation. Afrogarypus excelsus excellens is thus synonymised with A. excelsus stat. nov.
Additional material examined
SOUTH AFRICA: Eastern Cape: 5♀, 2♂, 2 Deutonymphs, Fort Beaufort, Fort Fordyce Nature Reserve, Afromontane forest, 32o 41’S, 26o 29’E, 1090 m a.s.l., leg. J.A. Neethling & C. Luwes, 29.XI.2012 (NMBA P00190); 1♀, Grahamstown [33o 18’S, 26o 31’E], leg. J. Hewitt, VIII.1918 (AMG slide); 3♀, 4♂, 2 egg sacks, Grahamstown, Pine Plantation [33o 18’S, 26o 31’E], leg. C. Kewi, 2.XII.1918 (AMG slide); 1♀, King William’s Town, Pirie Forest Reserve [32o 43’S, 27o 16’E], leg. R. Godfrey, 27.VIII.1908 (AMG slide); 2♀, 2♂, Somerset East, Bestershoek Nature Reserve, Dry-shrub forest, 32o 42’S, 25o 33’E, 860 m a.s.l., leg. J.A. Neethling & C. Luwes, 2.XII.2012 (morphological dissection and S.E.M. material); 2♀, 2♂, Somerset East, Bosberg [32o 41’S, 25o 32’E], leg. R.F. Lawrence, III.1964 (NMSA 5126); 1♀, Somerset East, Zuurberg Pass, Afromontane forest clusters, 33o 17’S, 25o 42’E, 772 m a.s.l., leg. J.A. Neethling, 15.IX.2011 (phylogenetic analysis); 13♂, Stutterheim, Sandile’s Rest Lodge, Afromontane forest, 32o 39’S, 27o 17’E, 902 m a.s.l., leg. J.A. Neethling, 20.IV.2013 (NMBA P00191).
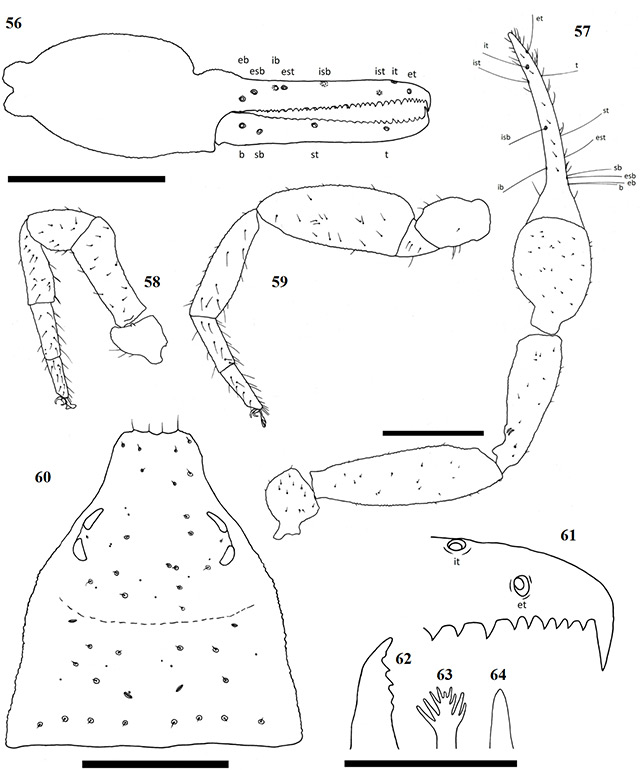
Figures 56–64. Afrogarypus excelsus (Beier, 1964) stat. nov. 56–63. Female; 64. Male. 56. Right chela, retrolateral view; 57. Right pedipalp, dorsal view; 58. Right leg I, prolateral view; 59. Right leg IV, prolateral view; 60. Carapace, dorsal view; 61. Right chela fixed finger, anterior teeth arrangement; 62. Chelicera movable finger, dorsal view; 63, 64. Galea. Scale bars: Figs 56-60: 0.50 mm; Figs 61-64: 0.10 mm.
Additional material not examined
SOUTH AFRICA: Eastern Cape: 3♀, 3♂, Somerset East, Bosberg [32o 41’S, 25o 32’E], III.1964 (examined by Beier 1966).
Afrogarypus impressus (Tullgren, 1907)
Figs 65-77
Garypus impressus Tullgren, 1907b: 228, fig. 7 (Holotype ♀, South Africa, KwaZulu-Natal,
Amanzimtoti [30o03’S, 30o53’E], leg. I. Trägårdh, 18.I.1905, collection unknown, not examined; Paratypes, South Africa, KwaZulu-Natal, Hluhluwe-Imfolozi Game Reserve, Junction of Black and White Imfolozi Rivers [28o13’S, 31o57’E], leg. I. Trägårdh, 28.VIII.1905, collection unknown, not examined; Van Reenen, [28o22’S, 29o23’E], leg. I. Trägårdh, 24.VI.1905, collection unknown, not examined); Ellingsen, 1912: 86, 107.
Geogarypus impressus (Tullgren, 1907): Chamberlin, 1930: 609; Roewer, 1936: fig. 77;
Beier, 1964: 60; Lawrence, 1967: 89; Spaull, 1979: 117.
Geogarypus (Afrogarypus) impressus (Tullgren, 1907): Beier, 1932: 236, fig. 265; Roewer,
1937: 270, fig. 222b; Beier, 1947: 320; Beier, 1955: 301, fig. 21a; Beier, 1958: 171; Beier, 1966: 461.
Afrogarypus impressus (Tullgren, 1907): Harvey, 1986: 758; Harvey, 1991: 250; Dippenaar- Schoeman & Harvey, 2000: 93.
Diagnosis
Large species. Females reaching a total length (cucullus to posterior abdominal margin) of 3.25 mm, with males reaching up to 2.84 mm. Carapace dark brown, medial furrow and posterior margin light brown. All pedipalp segments dark brown in colour. Well developed sulcus present on dorsal surface of chela. Abdominal tergites mostly medium brown with dark patches, abdominal sternites pale and weakly sclerotised anteriorly to medium brown and well sclerotised posteriorly (♀), brown with indistinct, paired, darker patches and well sclerotised throughout (♂). Pedipalp coxae brown in colour with distinct shoulder, legs I–IV as well as remaining coxae tan to pale yellow. Both sexes are quite variable in size, though males still tend to be smaller than the females from the same locality.
Closely resembling A. excelsus stat. nov. in both size and colouration, the two species are the only large South African geogarypids that possess a deep, well developed dorsal sulcus on each chela. Afrogarypus impressus can reliably be differentiated from A. excelsus stat. nov. by the ratios of its fingers to the hand-stem complex, where the fingers are longer than the complex in A. excelsus stat. nov. and shorter than the complex in A. impressus. Further differences include a broader and shorter carapace in A. impressus, the front four teeth on the fixed finger grouping on a bulge, as well as chelae that are broader compared to their length than those found in A. excelsus stat. nov. The anterior tergites of A. impressus furthermore possess distinct cream patches that are absent in A. excelsus stat. nov.
Description
Carapace: Strongly sub-triangular, with narrow furrow posterior to the eyes (Fig. 73). Uniformly dark brown, medial furrow and posterior margin light brown. Uniformly granular, heavily constricted anteriorly into cucullus, constriction beginning between medial furrow and posterior pair of eyes. Two pairs of corneate eyes situated on ocular tubercles, located about one-third away from the anterior edge. Four prominent setae located on anterior edge; row of eight setae, seated within large rims, located on the posterior margin. Numerous small setae present on carapace, some situated within large rims.
Abdomen: Abdominal tergites brown (♀), light brown (♂), with dark patches. Tergites I and II with large median dark spot each. Tergite III without dark patches. Tergites IV–X with paired dark patches just lateral of midline, may be just as dark, or slightly lighter, than those on tergites I and II. Tergites XI and XII without patches. Sternites pale, weakly sclerotised anteriorly, brown and well sclerotised posteriorly (♀). Brown, with indistinct, paired darker patches, well sclerotised throughout (♂). Pleural membrane wrinkled-plicate with invested setae.
Pedipalp: Trochanter rounded, somewhat square in shape, granular, uniformly dark brown. Small setae scattered over entire surface, with exception of stem. Apophysis located ventrally. Pedipalp femur (Fig. 70) dark brown, granular, narrow at base. Widens evenly, interior margin then constricts anteriorly to form narrow end. Pedipalp patella (Fig. 70) dark brown and granular. Narrow at base, widening quickly into cone. Lyriform fissures visible just anterior of base, located on bulge, dorso-prolaterally on patella.
Chela: (Figs 65-70) Uniformly brown, granular from stem to base of fingers. Both fingers smooth. Well developed dorsal sulcus located just posterior of base of fixed finger. Dorsal bulge located at base of fixed chela finger. Fixed chela finger with eight trichobothria, movable chela finger with four, both fingers narrow, curving to interior. Fingers shorter than hand (with pedicel). Venom apparatus present on both fingers. Trichobothria eb and esb located retrolaterally, at base of fixed chela finger, situated ca. one trichobothrial width from each other; ib situated dorso-prolaterally, just distally of esb; est opposite and just anterior of ib; isb located pro-dorsally, aligned vertically with st, grouping close to est; ist located ca. one third from anterior edge of fixed finger, situated prolaterally; it located one quarter finger length from anterior edge, situated dorsally; et located distally from it, almost equidistant from it as ist from it. Regarding the movable finger, b located just anterior of finger base, situated dorso-retrolaterally on the finger in line vertically with eb; sb is located just anterior of b, situated slightly more ventrally on the finger, mostly aligned with esb; st is located ca. one third along movable finger, closer to sb than t; t located ca. one third from the anterior edge of the movable finger, just forward of ist. Chelal teeth acute and retrorse, some curving back slightly. Fixed finger with 31 teeth (♀), 25 (♂). Four closely spaced teeth, arranged on a slight bulge, just behind venom apparatus, followed by somewhat larger tooth (Fig. 74). Further teeth arranged, roughly, in two rows and reduce in size proximally, but remain acute. Movable finger with 17 teeth (♀), 6 (♂). In the female, movable finger’s teeth are nearly contiguous, reduced to small projections posteriorly after first seven teeth. In the male, only six well defined teeth visible posterior of the venom apparatus.
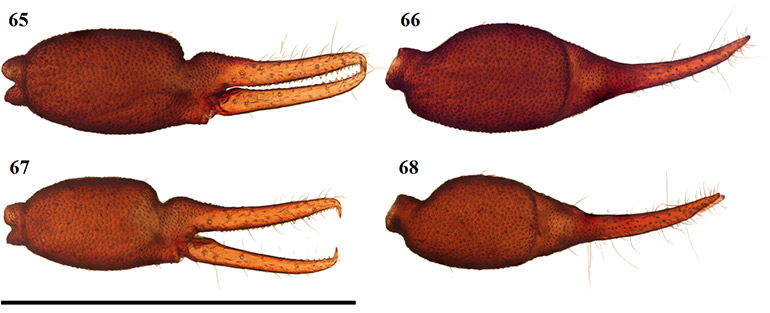
Figures 65–68. Digital microscope photographs of Afrogarypus impressus (Tullgren, 1907) right chela: Female (65, 66) and Male (67, 68). 65, 67. Retrolateral view; 66, 68. Dorsal view. Scale bar: 1.00 mm.
Chelicera: Hand with five setae as in Fig. 1. Fixed cheliceral finger with five teeth arranged as in Fig. 75. Galea complex, with nine rami arranged closer together on one side as in Fig. 76 (♀), simple, with no rami (♂) (Fig. 77). Rallum composed of single blade in both male and female. Serrula exterior with 17 lamella (♀) or 18 (♂). Lamina exterior present in both sexes.
Coxae and legs: Pedipalp coxae brown, shoulder distinct. Multiple long setae present along inner medial ridges. Coxae of legs I–IV, as well as all leg segments, tan. Trochanters of legs I and II compact and small, larger and elongate on legs III and IV. Femora of legs I and II longer than patellae and tibiae respectively (Fig. 71). Metatarsi and tarsi not fused, joint movable. Tarsal claws simple and curved, arolium much longer than claws, not divided. Femur-patella joint immovable in leg IV, tibia mostly straight (Fig. 72), metatarsi and tarsi not fused, joint movable. Claws and arolium similar to those of legs I–III.
Measurements: body length ♀ 3.25 ♂ 2.84; carapace ♀ 1.16 x 1.22 (0.95) ♂ 0.95 x 0.97 (0.98). Palp femur ♀ 1.26 x 0.38 (3.31) ♂ 1.04 x 0.32 (3.25), patella ♀ 0.98 x 0.34 (2.88) ♂ 0.81 x 0.30 (2.70), hand ♀ 0.65 x 0.34 ♂ 0.55 x 0.30, chela ♀ 1.14 (3.35) ♂ 1.06 (3.53), movable finger ♀ 0.48 ♂ 0.49. Leg I femur ♀ 0.51 x 0.20 ♂ 0.46 x 0.19, patella ♀ 0.28 x 0.18 ♂ 0.25 x 0.18, tibia ♀ 0.34 x 0.14 ♂ 0.32 x 0.13, metatarsus ♀ 0.24 x 0.11 ♂ 0.23 x 0.10, tarsus ♀ 0.24 x 0.06 ♂ 0.22 x 0.07. Leg IV femur ♀ 0.29 x 0.18 ♂ 0.25 x 0.15, patella ♀ 0.82 x 0.28 ♂ 0.67 x 0.28, tibia ♀ 0.68 x 0.17 ♂ 0.53 x 0.16, metatarsus ♀ 0.37 x 0.12 ♂ 0.29 x 0.11, tarsus ♀ 0.32 x 0.08 ♂ 0.26 x 0.08.
Remarks
This is one of the first three geogarypid species to be described from South Africa, along with A. minutus comb. nov. and G. olivaceus. Tullgren (1907b) failed to mention where he lodged the type specimens. It was assumed that they would reside in ZMH along with the types for A. minutus comb. nov. and G. olivaceus, but this is not the case. Beier (1932) gave a short description of specimens he studied that were lodged in the Museum für Naturkunde of Berlin.
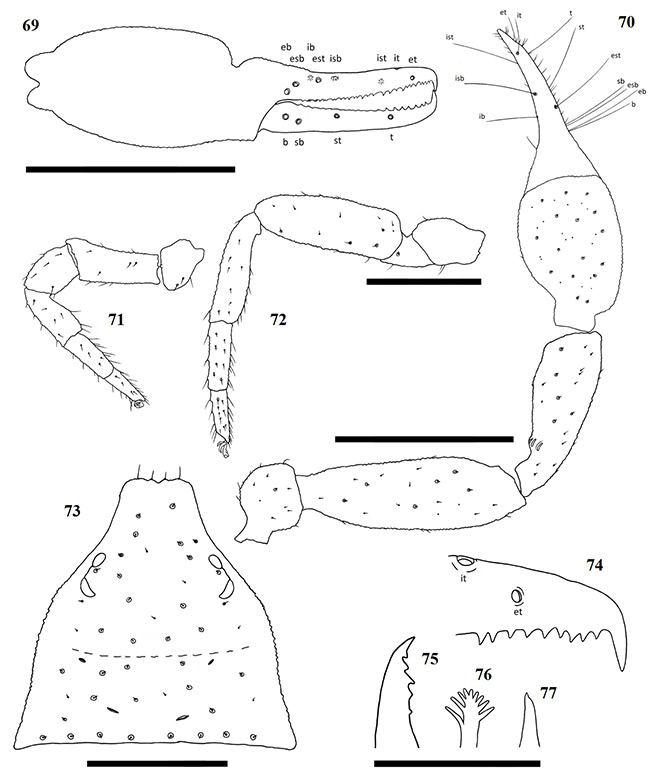
Figures 69–77. Afrogarypus impressus (Tullgren, 1907). 69–76. Female; 77. Male. 69. Right chela, retrolateral view; 70. Right pedipalp, dorsal view; 71. Right leg I, prolateral view; 72. Right leg IV, prolateral view; 73. Carapace, dorsal view; 74. Right chela fixed finger, anterior teeth arrangement; 75. Chelicera movable finger, dorsal view; 76, 77. Galea. Scale bars: Figs 69-73: 0.50 mm; Figs 74-77: 0.10 mm.
Additional material examined
SOUTH AFRICA: Eastern Cape: 1♀, Grahamstown, Paradise Kloof [33o 18’S, 26o 31’E], leg. J. Hewitt, I.1919 (AMG Slide); 1♂, Kei Mouth, coastal forest, 32o 41’S, 28o 22’E, 21 m a.s.l., leg. C.R. Haddad, 8.XII.2005 (WAM T79019); 1♀, 2 Deutonymphs, King William’s Town, Pirie Forest Reserve [32o 43’S, 27o 16’E], leg. R. Godfrey, IX.1913 (NMSA 16050); 2♂, Stormsrivier, Stormsrivier Forest Nature Reserve, indigenous forest, 33o 59’S, 23o 54’E, 185 m a.s.l., leg. J.A. Neethling, 5.XII.2012 (NMBA P00171); 1♀, 6♂, Stormsrivier, Tsitsikamma National Park, coastal forest, 34o 01’S, 23o 53’E, 47 m a.s.l., leg. J.A. Neethling & C. Luwes, 22.XII.2013 (NMBA P00170); 2♀, 3♂, Same locality, Stormsrivier Mouth, 34o 01’S, 23o 54’E, 39 m a.s.l. leg. J.A. Neethling & C. Luwes, 22.XII.2013 (morphological dissection); 2♂, 4 Tritonymphs, Same locality [34o 01’S, 23o 54’E], leg. R.F. Lawrence & J.Y. Lawrence, I.1961 (NMSA 7866). KwaZulu-Natal: 7♀, 3♂, Durban [29o 51’S, 31o 01’E], leg. W.G. Rump, VIII.1940 (NMSA 5132); 1 protonymph, Izindophi, Middle Drift, Tugela River [28o 53’S, 31o 01’E], leg. R.F. Lawrence, X.1940 (NMSA 5131); 1♂, Margate [30o 51’S, 30o 22’E], leg. W.G. Rump, IV.1940 (NMSA 5130); 18♀, 13♂, 1 deutonymph, 2 tritonymphs, Pietermaritzburg, Table Mountain [29o 36’S, 30o 36’E], leg. R.F. Lawrence, I.1940 (NMSA 5127); 1♀, 2♂, 1 Deutonymph, 2 Tritonymphs, Summerveld, Shongweni Resource Reserve [29o 50’S 30o 43’E], leg. W.G. Rump, VII.1940 (NMSA 5128). Western Cape: 1♀, George, Groenkop, Groeneweide Forest [33o 56’S, 22o 32’E], leg. J.H. Koen, VII.1986 (NCA 97/313); 1♀, 1♂, George, Saasveld Pass, indigenous forest, 33o 58’S, 22o 31’E, 149 m a.s.l., leg. J.A. Neethling, 7.XII.2012 (phylogenetic analysis); 1♀, George, Wadeville Forest, indigenous evergreen forest, 33o 55’S, 22o 39’E, 380 m a.s.l., leg. H.G. Robertson, 13.X.1996 (SAMC ENW-COO 5420); 1♀, 1♂, Hoekwil, Woodville Big Tree, 33o 56’S, 22o 38’E, 262 m a.s.l., leg. J.A. Neethling, 8.XII.2012 (lactic acid clearing); 1♀, Knysna, Diepwalle Forest, indigenous forest [33o 57’S, 23o 09’E], leg. J.H. Koen, III.1984 (NCA 97/281); 2♀, Knysna, Lily Vlei Nature Reserve, Gouna State Forest, indigenous forest [33o 56’S, 23o 02’E], leg. J.H. Koen, VII.1983 (NCA 97/371); 1♀, 2♂, Mossel Bay, St. Blaize Hiking Trail, coastal fynbos, 34o 11’S, 22o 08’E, 89 m a.s.l., leg. J.A. Neethling, 8.XII.2012 (NMBA P00169); 2♀, 2♂, Rheenendal, Jubilee Creek Nature Reserve, 33o 53’S, 22o 58’E, Afromontane forest, 923 m a.s.l., leg. J.A. Neethling & C. Luwes, 26.XII.2013 (NMBA P00172); 1♀, Suurbraak, Grootvadersbosch Nature Reserve, dry shrub forest, 33o 58’S, 20o 49’E, 422 m a.s.l., leg. J.A. Neethling, 13.XII.2012 (phylogenetic analysis).
Additional material not examined
SOUTH AFRICA: KwaZulu-Natal: 1♂, Royal Natal National Park, Giants Castle Game Reserve, Mahai Rest Camp, The Cascades [28o 41’S, 28o 56’E], 2.IV.1951 (examined by Beier 1955, collection unknown); St. Lucia, iSimangaliso Nature Reserve, Lake Sibayi shoreline [27o 21’S, 32o 43’E] (examined by Beier 1958, collection unknown).
Northern Cape: 2 immatures, Steinkopf [29o 15’S, 17o 44’E], leg. Schultze (examined by Beier 1947, deposited in ZMH).
Afrogarypus megamolaris sp. nov.
Figs 78–94
Holotype: ♀, SOUTH AFRICA, Limpopo Province, Makhado [Louis Trichardt], Medike Mountain Reserve, 22o 59’S, 29o 37’E, 903 m a.s.l., Montane Bushveld, Leaf litter sifting, leg. J.A. Neethling, 9.XI.2012 (NMBA P00206).
Paratype: 1♂, Same data as holotype (NMBA P00207).
Etymology
Named for the enlarged basal teeth on both chelal fingers of females.
Diagnosis
Small species. Females can reach up to 1.80 mm and males up to 1.53 mm in total length (cucullus to posterior abdominal margin). Carapace uniform brown to dark brown in both sexes, though the medial furrow and posterior margin may be slightly lighter in colour than the rest of the carapace. All pedipalp segments similar in colouration to the carapace. Well developed, broad and shallow sulcus present on the dorsal surface of chela. Abdominal tergites generally medium brown with dark patches, though small cream coloured markings may be present on each side of the paired dark spots. Abdominal sternites light brown with cream markings in both sexes, weakly sclerotised in females and strongly sclerotised throughout in males. Pedipalp coxae has same colouration as carapace and possesses a distinct shoulder. Legs I–IV, as well as remaining coxae, tan to pale yellow.
This species resembles A. minutus comb. nov. in general size and appearance, but can be easily distinguished by the presence of a well developed dorsal sulcus on the chela hand, while A. minutus comb. nov. possesses only a shallow concave area.
Description
Carapace: Strongly sub-triangular, narrow furrow posterior to the eyes (Fig. 92). Uniformly medium brown to dark brown in both sexes, medial furrow and posterior margin somewhat lighter. Uniformly granular, heavily constricted anteriorly into cucullus, constriction beginning at the medial furrow. Row of eight setae located on the posterior edge. Two pairs of corneate eyes situated on ocular tubercles, located about one-third away from the anterior edge.
Abdomen: Abdominal tergites uniformly medium brown, with dark patches (small cream patches may be present, flanking paired dark spots) in both males and females, granular. Tergites I and II each with large median dark spot. Tergite III without dark patches. Tergites IV–X with paired, indistinct dark patches mediolaterally. Tergites XI and XII without patches. Sternites light brown with indistinct dark patches in both sexes, weakly sclerotised in females, well sclerotised in males. Pleural membrane wrinkled-plicate, cream.
Pedipalp: Trochanter rounded, compact in shape, granular, coloured same as carapace. Small setae scattered over entire surface, except the stem. Apophysis located ventrally. Pedipalp femur (Fig. 85) same colour, granular. Narrow at stem, widening quickly at base, widening further slightly before constricting again at the end. Pedipalp patella (Fig. 85) granular and same colour as femur. Narrow and slightly angled at base, widening evenly into cone. Lyriform fissures visible just anterior to base, located on a bulge, dorso-prolaterally on patella.
Chela: (Figs 78–81, 84, 85) Same colour as rest of pedipalp, granular from stem to base of fingers. Both fingers smooth, just as long to slightly longer than hand (with pedicel). Well developed, broad, shallow sulcus present on dorsal surface of chelal hand. Chela hand rounded at back on prolateral edge, tapering in toward base of fingers with a straight edge. Retrolateral edge mostly straight. Fixed chela finger with eight trichobothria, movable chela finger with four, both fingers narrow, curving slightly to interior. Venom apparatus present on both fingers. Trichobothria eb and esb situated as normal on retrolateral surface of fixed chelal finger, located just above base of finger, one trichobothrial width between them. Trichobothria ib located on the prolateral surface, anterior and dorsal of esb. Trichobothria est located opposite and anterior of ib by slightly more than one trichobothrial width. Trichobothria isb located just before halfway along fixed finger, situated dorsally, while ist located on interior lateral surface, over one-third from anterior edge of fixed chelal finger, situated near the teeth. Trichobothria it located dorsally, roughly one-quarter distance from anterior edge of fixed chelal finger, et located on retrolateral surface, slightly distal and almost beneath it. Trichobothria b, sb and st group together in triangle within the first third of movable finger, t located distally, almost directly in line with it. Chelal teeth acute, most slightly retrorse. Fixed finger with 27 teeth (♀), 30 (♂). First six teeth from the anterior end small, increasing slightly in size before the first large accessory tooth (Fig. 88). Further teeth slightly retrorse and acute, arranged, mostly, in single row. Movable finger with 21 teeth (♀), 22 (♂) tooth just before venom apparatus very small. Teeth after small tooth acute and retrorse, decreasing in size/reducing into small points closer to base of finger. In females, first six basal teeth on fixed chelal finger, and first three on movable finger very enlarged, grouping tightly next to each other (Figs 82 & 86). In males, teeth remain small and acute (Figs 83 & 87).
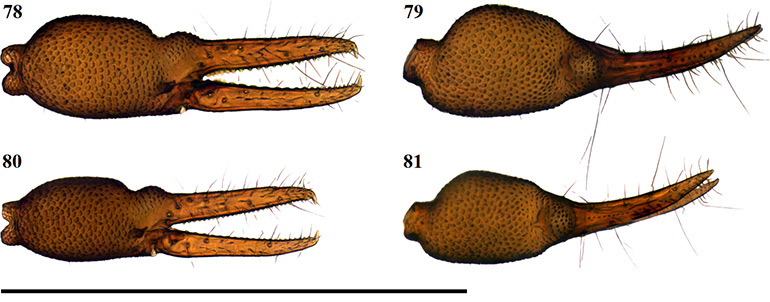
Figures 78–81. Digital microscope photographs of Afrogarypus megamolaris sp. nov. right chela: Female (78, 79) and Male (80, 81). 78, 80. Retrolateral view; 79, 81. Dorsal view. Scale bar: 1.00 mm.
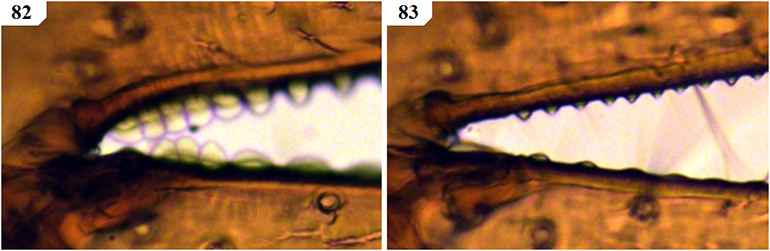
Figures 82-83. Tooth morphology at the base of the chelal fingers of Afrogarypus megamolaris sp. nov. Female (82) and Male (83), showing distinct differences between the genders.
Chelicera: Hand with five setae located as in Fig. 1. Fixed cheliceral finger with five teeth arranged as in Fig. 89. Female galea complex, with nine rami in a fan shape (Fig. 90), male galea simple, no rami (Fig. 91). Rallum composed of single blade in both sexes. Serrula exterior with 13 lamella (♀) or 12 (♂). Lamina exterior present in both sexes.
Coxae and legs: Pedipalp coxae light brown, shoulder distinct. Coxa of legs I–IV, as well as all leg segments, tan to pale yellow. Leg morphology standard and diplotarsate for both leg I (Fig. 94) and leg IV (Fig. 93).
Measurements: body length ♀ 1.80 ♂ 1.53; carapace ♀ 0.49 x 0.54 (0.90) ♂ 0.42 x 0.47 (0.89). Palp femur ♀ 0.46 x 0.16 (2.87) ♂ 0.44 x 0.14 (3.14), patella ♀ 0.40 x 0.15 (2.66) ♂ 0.35 x 0.14 (2.50), hand ♀ 0.38 x 0.24 ♂ 0.32 x 0.20, chela ♀ 0.77 (3.20) ♂ 0.68 (3.40), movable finger ♀ 0.38 ♂ 0.35. Leg I femur ♀ 0.21 x 0.09 ♂ 0.19 x 0.08, patella ♀ 0.12 x 0.08 ♂ 0.11 x 0.08, tibia ♀ 0.15 x 0.06 ♂ 0.13 x 0.06, metatarsus ♀ 0.12 x 0.04 ♂ 0.11 x 0.04, tarsus ♀ 0.11 x 0.03 ♂ 0.10 x 0.03. Leg IV femur ♀ 0.10 x 0.07 ♂ 0.10 x 0.06, patella ♀ 0.32 x 0.12 ♂ 0.28 x 0.11, tibia ♀ 0.30 x 0.07 ♂ 0.26 x 0.07, metatarsus ♀ 0.15 x 0.05 ♂ 0.13 x 0.05, tarsus ♀ 0.15 x 0.04 ♂ 0.13 x 0.03.
Remarks
Specimens of this species were primarily collected from the type locality, though museum specimens added further locality data. This species could have accounted for many A. impressus misidentifications within north-eastern areas of South Africa due to the presence of a dorsal sulcus, as such mis-identifications were present in museum samples (NMSA 16051) that corresponded to sample data in Beier (1966) and Lawrence (1967).
Additional material examined
SOUTH AFRICA: Gauteng Province: 2 adults, Pretoria, Groenkloof [25o 46’S, 28o 13’E], 15.IX.1921 (AMG slide); 1♀, Pretoria, Roodeplaat, Pretoria Plant Protection Research Institute [25o 36’S, 28o 21’E], leg. M.S. Harvey, 24.III.2001 (WAM T67019).
Mpumalanga Province: 1♀, Kruger National Park, Malelane Camp [25o 29’S, 31o 31’E], leg. R.F. Lawrence, 3.XII.1963 (NMSA 16051).
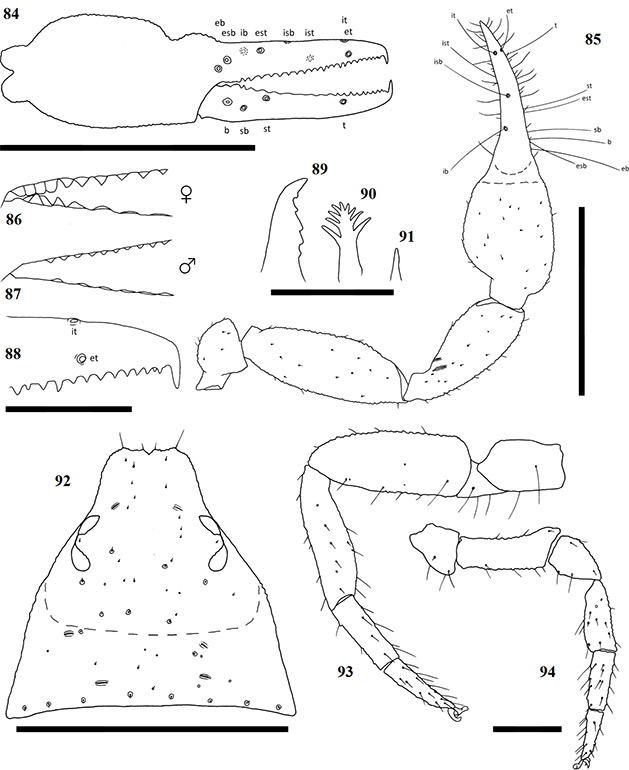
Figures 84–94. Afrogarypus megamolaris sp. nov. 84–86, 88–90, 92–94. Female; 87, 91. Male. 84. Right chela, retrolateral view; 85. Right pedipalp, dorsal view; 86, 87. Basal teeth arrangement; 88. Anterior teeth arrangement on fixed chelal finger; 89. Chelicera movable finger, dorsal view; 90, 91. Galea; 92. Carapace, dorsal view; 93. Right leg IV, prolateral view; 94. Right leg I, prolateral view. Scale bars: Figs 84, 85, 92: 0.50 mm; Figs 86-91, 93, 94: 0.10 mm.
Afrogarypus minutus (Tullgren, 1907) comb. nov.
Figs 95-109
Garypus minutus Tullgren, 1907a: 65, fig. 21 (Holotype ♀, South Africa, Eastern Cape, Port Elizabeth [33o 56’S, 25o 34’E], leg. H. Brauns, 15.XII.1898, deposited in ZMH,
examined); Tullgren, 1907b: 229; Ellingsen, 1912: 87, 108.
Geogarypus minutus (Tullgren, 1907): Chamberlin, 1930: 609; Beier, 1964: 60; Harvey,
1986: 760; Harvey, 1991: 257; Dippenaar-Schoeman & Harvey, 2000: 93; Turienzo, Di Iorio & Mahnert, 2010: 561, 564 [comb. nov.].
Geogarypus (Geogarypus) minutus (Tullgren, 1907): Beier, 1932: 231, fig. 257; Roewer,
1937: 269; Beier, 1958: 171.
Diagnosis
Small species. Females can reach a length (cucullus to posterior abdominal margin) of 1.60 mm, with males reaching 1.45 mm. Carapace dark brown throughout in both sexes, though in some specimens the medial furrow and posterior edge of the carapace may be lighter. Pedipalp trochanter, femur and patella light brown in colour, with pedipalp coxae and chela dark brown. Concave depression present on dorsal surface of chela. Abdominal tergites generally dark brown in females, dark brown with cream patches in males. Abdominal sternites cream to dark brown in both sexes, weakly sclerotised in females and well sclerotised in males. Pedipalp coxae possessing a distinct shoulder, legs I–IV cream in colour (both sexes), with the leg coxae yellow to light brown in females and cream in males.
Specimens closely resemble smaller individuals of A. triangularis comb. nov., but can be distinguished by the shape of the chela and the lack of lateral sclerites on the abdomens of males. The chela of A. minutus comb. nov. does not possess the strongly convex interior lateral surface that gives the chela of A. triangularis comb. nov. its distinct shape, and instead has a less convex shape. The prolateral surface of the chela of some specimens was found to be almost straight, giving the chela a distinct rectangular shape.
Description
Carapace: Strongly sub-triangular, narrow furrow posterior to the eyes (Fig. 107). Dark brown throughout in both sexes, medial furrow and posterior edge may be paler. Uniformly granular, strongly constricted anteriorly into cucullus, constriction beginning at the medial furrow. Two pairs of corneate eyes situated on ocular tubercles, located about one-third away from anterior edge. Four prominent setae located on anterior edge, row of seven setae located on posterior margin. Numerous small setae present on the carapace.
Abdomen: Abdominal tergites granular, uniformly dark brown in females, dark brown with cream patches in males. Tergites I and II with large median dark spot each. Tergite III without dark patches. Tergites IV–X with indistinct paired dark patches medio-laterally. Tergites XI and XII without patches. Sternites cream to dark brown in both sexes, weakly sclerotised in females, well sclerotised in males. Pleural membrane wrinkled-plicate, cream.
Pedipalp: Trochanter rounded in front, rectangular in overall shape, granular and uniformly light brown. Small setae scattered over entire surface, except the stem. Apophysis located ventrally. Pedipalp femur (Fig. 100) light brown, granular. Narrow at stem, widening quickly to form base, then widening evenly before constricting again at distal end. Pedipalp patella (Fig. 100) same colour as femur, granular. Narrow, angled at base, widening quickly into short cone. Lyriform fissures visible just anterior to base, located on bulge, dorso-prolaterally on patella.
Chela: (Figs 95–100) Uniformly dark brown, almost same colour as carapace, granular from stem to base of fingers. Both fingers smooth. Concave depression present on dorsal surface of chela, resulting in bulge at base of fixed finger. Chela hand somewhat convex on prolateral edge, retrolateral surface mostly straight. Prolateral surface of some specimens also almost straight, giving chela distinct rectangular shape. Fixed chela finger with eight trichobothria, movable chela finger with four, both fingers narrow, curving slightly to interior. Fingers slightly longer than hand (with pedicel). Venom apparatus present on both fingers. Trichobothria eb and esb located retrolaterally at base of fixed chela finger, situated about one trichobothrial width from each other. Trichobothria ib situated dorso-prolaterally, one trichobothrial width anterior of esb. Trichobothria est located retrolaterally, one trichobothrial width forward of ib. Trichobothria ist situated dorso-prolaterally just over one-third finger length from base of fixed finger. Trichobothria it located one-quarter finger length from anterior edge and situated dorsally; et located retrolaterally and distally from it. Regarding the movable finger, trichobothria b, sb and st located in first third of movable finger, just anterior to finger base and situated retrolaterally, b is furthermore situated in line vertically with eb. Trichobothria t located less than one-third from anterior edge of movable finger, positioned between ist and it. Chelal teeth acute and retrorse, some curving back slightly. Fixed finger with 29 teeth (♀), 28 (♂). First four to five teeth after venom apparatus small, roughly equal in size, situated just in front of first large accessory tooth (Fig. 103). Additional teeth slightly to distinctly retrorse and acute, arranged roughly in two rows. Movable finger with 20 teeth (♀), 22 (♂). In both sexes, first eight teeth following the venom apparatus acute and retrorse, the rest of the teeth decreasing in size into small tooth points. Six basal teeth in females increase in size (Fig. 101), as with the first three in males (Fig. 102).

Figures 95–98. Digital microscope photographs of Afrogarypus minutus (Tullgren, 1907) comb. nov. right chela: Female (95, 96) and Male (97, 98). 95, 97. Retrolateral view; 96, 98. Dorsal view. Scale bar: 1.00 mm.
Chelicera: Hand with five setae located as in Fig. 1. Fixed cheliceral finger with five teeth arranged as in Fig. 104. Female galea complex elongate with eight rami (Fig. 105), small, simple, with no rami in males (Fig. 106). Rallum composed of single blade in both sexes. Serrula exterior with 16 (♀) or 14 (♂) lamella. Lamina exterior present in both sexes.
Coxae and legs: Pedipalp coxae same colour as carapace, with distinct shoulder. Legs I–IV cream in both sexes, leg coxae yellow to light brown in females, cream in males. Leg morphology standard and diplotarsate with regard to both leg I (Fig. 109) and leg IV (Fig. 108).
Measurements: body length ♀ 1.60 ♂ 1.45; carapace ♀ 0.51x 0.49 (1.04) ♂ 0.47 x 0.43 (1.09). Palp femur ♀ 0.52 x 0.16 (3.25) ♂ 0.44 x 0.14 (3.14), patella ♀ 0.43 x 0.15 (2.86) ♂ 0.37 x 0.13 (2.84), hand ♀ 0.39 x 0.26 ♂ 0.34 x 0.23, chela ♀ 0.81 (3.11) ♂ 0.71 (3.08), movable finger ♀ 0.40 ♂ 0.34. Leg I femur ♀ 0.22 x 0.09 ♂ 0.20 x 0.08, patella ♀ 0.13 x 0.08 ♂ 0.11 x 0.07, tibia ♀ 0.17 x 0.07 ♂ 0.15 x 0.06, metatarsus ♀ 0.13 x 0.05 ♂ 0.10 x 0.04, tarsus ♀ 0.13 x 0.04 ♂ 0.11 x 0.03. Leg IV femur ♀ 0.11 x 0.08 ♂ 0.09 x 0.07, patella ♀ 0.33 x 0.13 ♂ 0.31 x 0.14, tibia ♀ 0.30 x 0.08 ♂ 0.29 x 0.08, metatarsus ♀ 0.16 x 0.05 ♂ 0.14 x 0.05, tarsus ♀ 0.15 x 0.04 ♂ 0.14 x 0.04.
Remarks
This is one of the first three geogarypid species described for South Africa by Tullgren (1907a). The species was originally the smallest of all South African geogarypids, though it now places second after Afrogarypus castigatus sp. nov. which has an average length of 1.40 mm. Afrogarypus minutus comb. nov. is here transferred to the Afrogarypus due, firstly, to its phylogenetic placement within the Afrogarypus clade and secondly, due to the presence of a concave dorsal depression on the chela hand, a feature it shares with other species originally placed within the Geogarypus but now grouped within Afrogarypus. This species is restricted to the Eastern Cape and eastern Western Cape.
Additional material examined
SOUTH AFRICA: Eastern Cape: 1♀, Graaff-Reinet, Asante Sana Game Reserve, Waterkloof, 32o 15’S, 24o 56’E, leg. J. Midgely, 25.IX.2009 (NCA 2012/3500); 2 Adults, Grahamstown [33o 18’S, 26o 31’E] (AMG slide); 1 Adult, Port Alfred [33o 36’S, 26o 53’E], leg. J. Hewitt (AMG slide); 2♀, 2♂, Port Elizabeth, Island Nature Reserve, 33o 59’S, 25o 21’E, 268 m a.s.l., Indigenous bush, Leaf litter sifting, leg. J.A. Neethling, 15.XII.2013 (NMBA P00186); 2 Adults, Tsolo, Somerville Mission Station [31o 12’S, 28o 39’E], leg. R. Godfrey, VI.1912 (AMG slide). Western Cape: 2♀, 1♂, Knysna, Brenton-on-Sea, 34o 04’S, 23o 00’E, 228 m a.s.l., Coastal Fynbos, Leaf litter sifting, leg. J.A. Neethling, 20.XII.2013 (NMBA P00185); 3♀, 1 protonymph, 2 deutonymphs, Knysna, Brenton-on-Sea [34o 04’S, 23o 00’E], leg. H.G. Robertson, 19-23.XI.1998 (SAMC ENW-COO 5846, 5848, 5855, 5856, 5859); 1♀, Mossel Bay [34o 11’S, 22o 08’E], leg. W.F. Purcell, 1896 (SAMC 588).
Additional material not examined
SOUTH AFRICA: KwaZulu-Natal: Durban, Stamfort Hill [29o 50’S, 31o 02’E], leg. I. Trägårdh, 31.I.1905 (examined by Tullgren 1907b, deposited in ZMH); Van Reenen [28o
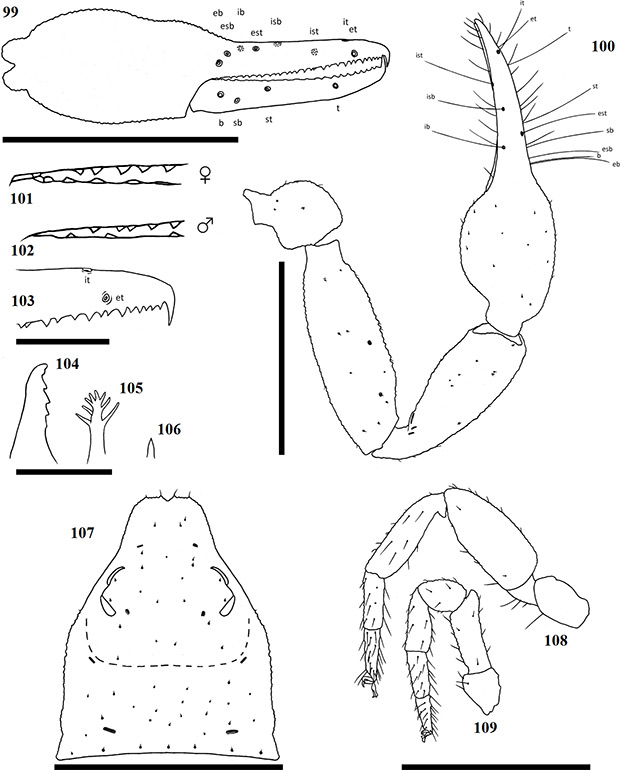
Figures 99–109. Afrogarypus minutus (Tullgren, 1907) comb. nov. 99–109, 103–105, 107–109. Female; 102, 106. Male. 99. Right chela, retrolateral view; 100. Right pedipalp, dorsal view; 101, 102. Basal teeth arrangement; 103. Anterior teeth arrangement on fixed chelal finger; 104. Chelicera movable finger, dorsal view; 105, 106. Galea; 107. Carapace, dorsal view; 108. Right leg IV, prolateral view; 109. Right leg I, prolateral view. Scale bars: Figs 99, 100, 107–109: 0.50 mm; Fig. 103: 0.10 mm; Figs 104-106: 0.05 mm.
23’S, 29o 23’E], leg. I. Trägårdh, 23.IV.1905 (examined by Tullgren 1907b, deposited in ZMH). Western Cape: 2♀, Caledon [34o 14’S, 19o 26’E], leg. W.F. Purcell (examined by Ellingsen 1912); 1♀, Clanwilliam, Waterfall Kloof [32o 04’S, 19o 05’E], leg. R. Pattison (examined by Ellingsen 1912); 1 immature, George, Montagu Pass [33o 54’S, 22o 25’E], leg. W.F. Purcell (examined by Ellingsen 1912); 1♂, Robertson [33o 48’S, 19o 53’E], leg. W.F. Purcell (examined by Ellingsen 1912); 1♀, Swellendam, Zondereinde Mountains [34o 01’S, 20o 26E’], leg. W.F. Purcell (examined by Ellingsen 1912).
Afrogarypus purcelli (Ellingsen, 1912) comb. nov.
Figs 110–122
Garypus purcelli Ellingsen, 1912: 110 (Holotype ♀, 1 immature, South Africa, Western
Cape, Beaufort West [32o 21’S, 22o 35’E], leg. W.F. Purcell, collection unknown, not examined).
Geogarypus (Geogarypus) purcelli (Ellingsen, 1912): Beier, 1932: 235; Roewer, 1937: 270;
Beier, 1955: 300, fig. 19.
Geogarypus purcelli (Ellingsen, 1912): Beier, 1964: 59; Beier, 1966: 461; Harvey, 1986:
760; Harvey, 1991: 259; Dippenaar-Schoeman & Harvey, 2000: 93 [comb. nov.].
Diagnosis
Medium to large species. Females can reach a length (cucullus to posterior abdominal margin) of 2.22 mm, with males reaching 2.14 mm. Carapace uniform medium brown to light brown in both sexes. Pedipalp segments same colour as carapace, while chela may be slightly darker. Concave depression present on dorsal surface of chela, with a distinct inclusion present on the prolateral surface just before the base of the fingers. Abdominal tergites mostly medium brown, with dark patches and a medial cream patch on most tergites, abdominal sternites brown and relatively well sclerotised (♀), well sclerotised throughout (♂). Pedipalp coxae same colour as carapace, with distinct shoulder, legs I–IV cream in colouration (both sexes), with leg coxae pale yellow to light brown.
Although somewhat larger, A. purcelli comb. nov. resembles A. carmenae sp. nov. in general morphology. The two can be distinguished by either the presence (A. purcelli comb. nov.) or lack (A. carmenae sp. nov.) of the inclusion on the prolateral surface of the chela. Mature females are generally larger and more robust than males, which are more elongate.
Description
Carapace: Strongly sub-triangular, longer than broad in both sexes, narrow furrow posterior to the eyes (Fig. 120). Carapace uniformly brown to light brown in both sexes, granular. Heavily constricted anteriorly into cucullus, constriction beginning at medial furrow. Two pairs of corneate eyes situated on ocular tubercles, located about one-third away from anterior edge. Four prominent setae located on anterior edge, row of eight setae, seated with rims, located on posterior margin. Numerous small setae present on carapace.
Abdomen: Abdominal tergites brown with dark patches and medial cream patch on most plates. Tergites I and II with large median dark spot each, flanked by cream patches. Tergite III without dark patches, but with cream patches. Tergites IV–X with indistinct paired dark patches mediolaterally. Tergites XI and XII lack dark patches. Tergites IV–IX with medial cream patch. Sternites relatively well sclerotised in females, well sclerotised in males. Brown to light brown throughout, with paired dark patches medio-laterally. Pleural membrane wrinkled-plicate, cream.
Pedipalp: Trochanter rounded, slightly elongate, granular, somewhat lighter in colour than carapace. Small setae located on dorsal and dorso-lateral sides. Apophysis located ventrally. Pedipalp femur (Fig. 115) and patella same colour as trochanter, granular. Femur narrow at base, widening evenly until just posterior of anterior margin, where there is a sudden constriction. Pedipalp patella (Fig. 115) narrow, slightly angled at base, widening evenly into elongate cone. Lyriform fissures visible just anterior of base, located on a bulge, dorso-prolaterally on patella.
Chela: (Figs 110-115) Slightly darker than rest of pedipalp in both sexes, granular from stem to base of fingers. Both fingers smooth, distinctly longer than hand (with pedicel). Dorsal depression present on chelal hand, resulting in bulge at base of fixed chelal finger. Prolateral surface slightly convex, ending in sharp inclusion at the base of chelal fingers, distinct to species. Fixed chela finger with eight trichobothria, movable chela finger with four. Both fingers long and narrow, curving to interior. Venom apparatus present on both fingers. Venom apparatus on female movable finger extends slightly past anterior edge of fixed chelal finger (Fig. 114). Trichobothria eb and esb situated just over one trichobothrial width from each other, located retrolaterally at base of fixed finger, while ib is located dorso-prolaterally, three trichobothrial widths distal of esb. Trichobothria est is located just short of one-third finger length on retrolateral surface, with isb situated distally by three trichobothrial widths on opposite surface. Trichobothria ist located one-third finger length from anterior finger edge, roughly in middle of prolateral surface; it located on dorsal surface, one-sixth finger length from anterior finger edge, while et is situated just anterior on retrolateral surface. Four trichobothria on the movable finger positioned with b ventrally in line, with eb and sb ventrally in line with ib. Trichobothria st positioned in line ventrally with isb, while t located roughly one-quarter finger length from anterior edge of the movable finger. Chelal teeth acute and retrorse. Fixed finger with 43 teeth (♀), 39 (♂). Eight to nine small, closely-spaced teeth just behind venom apparatus, followed by large accessory tooth (Fig. 116). Further teeth arranged roughly in two rows, decreasing in size to base of finger. Movable finger with 29 teeth (♀), 25 (♂). First tooth behind venom apparatus small, followed by large teeth that decrease in size into small points. In females, first five basal teeth increase in size into acute and retrorse structures, basal three spaced tightly next to each other. Slight teeth size increase in males.
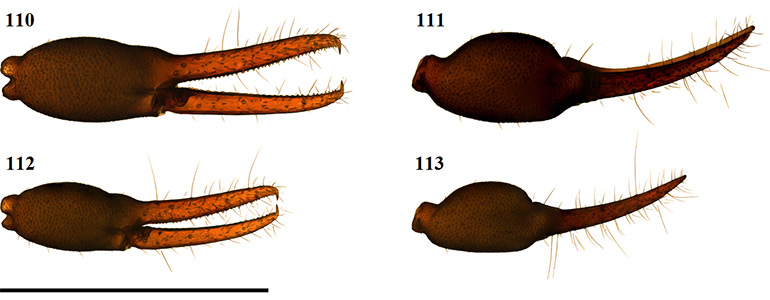
Figures 110–113. Digital microscope photographs of Afrogarypus purcelli (Ellingsen, 1912) comb. nov. right chela: Female (110, 111) and Male (112, 113). 110, 112. Retrolateral view; 111, 113. Dorsal view. Scale bar: 1.00 mm.
Chelicera: Hand with five setae located as in Fig. 1. Fixed cheliceral finger with five teeth arranged as in Fig. 117. Female galea complex, with nine rami (Fig. 118). Male galea simple, with no rami (Fig. 119). Rallum composed of single blade in both sexes. Serrula exterior with 19 lamella (♀) or 18 (♂). Lamina exterior present in both sexes.
Coxae and legs: Pedipalp coxae same colour as trochanters, with distinct shoulder. Legs I–IV cream in both sexes, with leg coxae yellow to light brown. Leg morphology standard and diplotarsate with regard to both leg I (Fig. 122) and leg IV (Fig. 121).
Measurements: body length ♀ 2.22 ♂ 2.14; carapace ♀ 0.79 x 0.70 (1.12) ♂ 0.64 x 0.54 (1.18). Palp femur ♀ 0.77 x 0.24 (3.20) ♂ 0.63 x 0.19 (3.31), patella ♀ 0.69 x 0.24 (2.87) ♂ 0.57 x 0.19 (3.00), hand ♀ 0.61 x 0.35 ♂ 0.48 x 0.27, chela ♀ 1.33 (3.80) ♂ 1.07 (3.96), movable finger ♀ 0.70 ♂ 0.58. Leg I femur ♀ 0.34 x 0.12 ♂ 0.29 x 0.10, patella ♀ 0.17 x 0.10 ♂ 0.16 x 0.10, tibia ♀ 0.26 x 0.09 ♂ 0.25 x 0.08, metatarsus ♀ 0.20 x 0.06 ♂ 0.16 x 0.06, tarsus ♀ 0.17 x 0.04 ♂ 0.16 x 0.04. Leg IV femur ♀ 0.16 x 0.11 ♂ 0.16 x 0.09, patella ♀ 0.53 x 0.19 ♂ 0.44 x 0.16, tibia ♀ 0.43 x 0.10 ♂ 0.38 x 0.10, metatarsus ♀ 0.23 x 0.07 ♂ 0.19 x 0.07, tarsus ♀ 0.22 x 0.05 ♂ 0.18 x 0.05.
Remarks
Afrogarypus purcelli comb. nov. is transferred to this genus due to the presence of a concave dorsal depression on the chelal hand, a character shared by A. robustus comb. nov., A. minutus comb. nov. and A. triangularis comb. nov. These species, originally placed in Geogarypus, group within the Afrogarypus clade in the molecular phylogeny, supporting their transfer. According to Ellingsen’s (1912) original description, type material for this species was housed with that of A. triangularis comb. nov. in the SAMC, although the types of neither species could be located during a visit to the museum. The outlying record from Makhado (Louis Trichardt) in Limpopo could represent a mis-identification, as no specimens belonging to the species were found during field work at this locality. Since no specimens were collected from the type locality, all identifications are regarded as tentative until new material is available.
Additional material examined
SOUTH AFRICA: KwaZulu-Natal: 1♀, Emangusi, Tembe Elephant Park, 26o 58’S, 32o 24’E, 112 m a.s.l., Sand forest, leg. C.R. Haddad, 12.IV.2006 (WAM T81144). Western Cape: 1♀, Arniston, Marcus Bay [34o 39’S, 20o 14’E], leg. H.A. Fry, 1900 (SAMC 13992); 8♂, Bredasdorp, De Hoop Nature Reserve, 34o 29’S, 20o 31’E, 26 m a.s.l., Coastal Fynbos, Leaf litter sifting, leg. J.A. Neethling & C. Luwes, 24.XII.2013 (NMBA P00189); 1♂, Bredasdorp, De Hoop Nature Reserve, Koppie Alleen, 34o 28’S, 20o 30’E, 53 m a.s.l., Rocky shore, leg. C.R. Haddad, 10.IV.2004 (WAM T79012); 1♂, Bredasdorp, De Hoop Nature Reserve, Lekkerwater Road, 34o 24’S, 20o
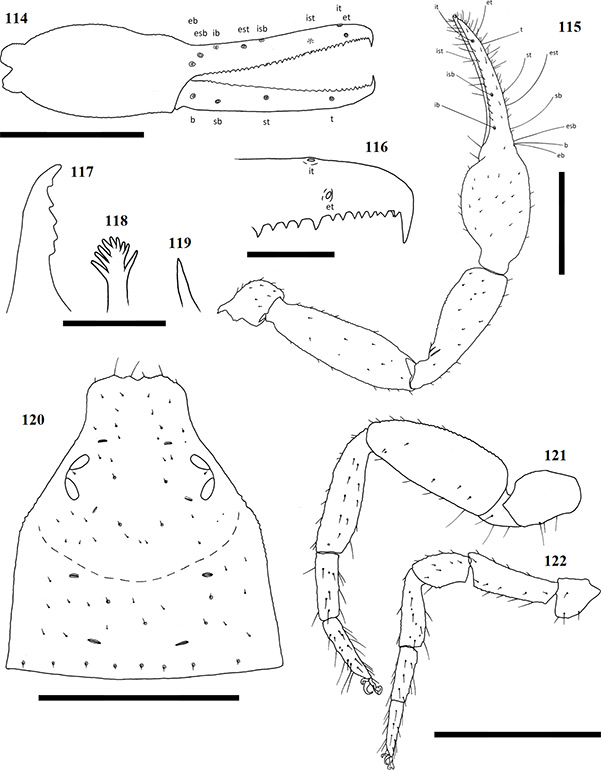
Figures 114–122. Afrogarypus purcelli (Ellingsen, 1912) comb. nov. 114–118, 120–122. Female; 119. Male. 114. Right chela, retrolateral view; 115. Right pedipalp, dorsal view; 116. Right chela fixed finger, anterior teeth arrangement; 117. Chelicera movable finger, dorsal view; 118, 119. Galea; 120. Carapace, dorsal view; 121. Right leg IV, prolateral view; 122. Right leg I, prolateral view. Scale bars: Figs 114, 115, 120-122: 0.50 mm; Fig. 116: 0.10 mm; Figs 117-119: 0.05 mm.
33’E, 134 m a.s.l., Grassland, leg. C.R. Haddad, 26.IX.2007 (WAM T86767); 2♂, Bredasdorp, De Hoop Nature Reserve, Potberg, 34o 22’S, 20o 32’E, leg. C.R. Haddad, 7.IV.2004 (WAM T79013).
Additional material not examined
SOUTH AFRICA: Limpopo Province: 93♀+♂, Makhado [Louis Trichardt], Mountain Top [23o 03’S, 29o 54’E], leg. R.F. Lawrence, II.1960 (examined by Beier 1964, deposited in NMZA). Western Cape: 1♂, Bredasdorp, 7 Miles SW [34o 36’S, 19o 57’E], 30.XII.1950 (examined by Beier 1955); 1♀, Bredasdorp, De Hoop Nature Reserve, 20 Miles ENE, Windhoek farm [34o 27’S, 20o 24’E], 2.I.1951 (examined by Beier 1955).
Afrogarypus robustus (Beier, 1947) comb. nov.
Figs 123–135
Geogarypus (Geogarypus) robustus Beier, 1947: 319, fig. 25 (Lectotype ♀ and
paralectotype, ♂, here designated, South Africa, Western Cape, Cape Town, Newlands, Fernwood [33o 59’S, 18o 27’E], leg. R.F. Lawrence (SAMC B8519), examined; Paratype 1♀, South Africa, Western Cape, Cape Town, St James [34o 07’S, 18o 27’E], leg. R.F. Lawrence (SAMC B8518), examined); Beier, 1955: 301.
Geogarypus robustus (Beier, 1947): Harvey, 1986: 760; Harvey, 1991: 259, Dippenaar-
Schoeman & Harvey, 2000: 93 [comb. nov.].
Diagnosis
Large species. Females can reach a length (cucullus to posterior abdominal margin) of 2.34 mm, with males reaching 2.23 mm. Carapace very dark, almost black, in females with medial furrow and posterior margin somewhat lighter in colour, while the male carapace is brown to red-brown. Female pedipalps dark in colouration, with chela slightly lighter, male pedipalps are the same colour as the carapace. Concave depression present on dorsal surface of chela. Abdominal tergites yellow to light brown with dark patches in females, red-brown with dark patches and a medial cream patch in males. Sternites well sclerotised in both sexes, yellow to light brown throughout in females, red-brown with dark patches and a cream-coloured medial line in males. Pedipalp coxae same colour as carapace with a distinct shoulder, legs I–IV grey in females, cream to dark brown in males.
Afrogarypus robustus comb. nov. is unique morphologically, with the abdomen being especially wide. Although easily distinguishable from all other South African Geogarypidae under a microscope, they do superficially resemble A. impressus to the naked eye, but lack a well developed sulcus on the chelal hand. The species is the most sexually dimorphic geogarypid in South Africa with regard to colouration, with females taking on a two-tone appearance (dark brown to black carapace and pedipalps, light coloured abdomen and legs) and males being more uniformly brown.
Description
Carapace: Strongly triangular in females and sub-triangular in males, with narrow furrow posterior to the eyes (Fig. 133). Much broader than long in females, with carapace of males being either just as broad as long or slightly longer than broad. Very dark to almost black, with slightly lighter furrow and posterior carapace margin in females. Uniformly brown to red-brown in males. Uniformly granular in both sexes, heavily constricted anteriorly into cucullus, constriction beginning at medial furrow. Two pairs of corneate eyes situated on ocular tubercles, located about one-third away from anterior edge. Four prominent setae located on anterior edge, row of nine setae, seated with rims, located on posterior margin. Numerous small setae present on carapace, some situated within rims.
Abdomen: Abdominal tergites yellow to light brown with dark patches in females. Red-brown with dark patches and median cream patch in males. Tergites I and II with large median dark spot each. Tergite III without dark patches. Tergites IV–X with paired dark patches just lateral of midline, while tergites XI and XII lack patches. Setae of posterior setal rows of tergites in both sexes situated on large pale plates with dark edges, making these plates distinctly visible. Sternites well sclerotised in both sexes, yellow to light brown throughout (♀), while red-brown with dark patches and cream midline (♂). Pleural membrane wrinkled-plicate, cream.
Pedipalp: Trochanter rounded, granular, slightly lighter than carapace. Small setae located on the dorsal and dorso-lateral sides. Apophysis located ventrally. Pedipalp femur (Fig. 128) same colour as trochanter, granular, narrow at base, widening evenly until just posterior of anterior margin where there is a sudden constriction. Pedipalp patella (Fig. 128) same colour and granulation as femur, narrow, slightly angled at base, widening evenly into elongate cone. Lyriform fissures visible just anterior of base, located on bulge, dorso-prolaterally on patella.
Chela: (Figs 123–128) Somewhat lighter than rest of pedipalp in females, same colour as rest of pedipalp in males. Both sexes granular from stem to base of fingers. Both fingers smooth, distinctly shorter than hand(with pedicel). Dorsal depression present on chelal hand, resulting in bulge at base of fixed chelal finger. Fixed chela finger with eight trichobothria, movable chela finger with four. Both fingers long and narrow, curving to interior. Venom apparatus present on both fingers. Venom apparatus on female movable finger extends slightly past anterior edge of fixed chelal finger (Fig. 127).

Figures 123–126. Digital microscope photographs of Afrogarypus robustus (Beier, 1947) comb. nov. right chela: Female (123, 124) and Male (125, 126). 123, 125. Retrolateral view; 124, 126. Dorsal view. Scale bar: 1.00 mm.
Trichobothria eb and esb situated just over one trichobothrial width from each other, located retrolaterally at base of fixed finger. Trichobothria est located retrolaterally within first third of fixed finger length, distal of esb, ib is situated almost exactly opposite, slightly more anterior on prolateral surface of finger. Trichobothria isb located roughly halfway along dorsal surface of finger, situated slightly prolaterally; ist located one-third finger length from anterior finger edge, situated in middle of prolateral surface. Trichobothria it located approximately one-quarter finger length from finger’s anterior edge, situated on dorsal surface, et located anteriorly on retro-dorsal surface. Regarding the movable finger, b and sb are located on retrolateral surface, just anterior of finger base, separated by two trichobothrial widths. Trichobothria st located just short of halfway along movable finger. Trichobothria t located one-quarter finger length from anterior finger edge, situated in line ventrally with it. Chelal teeth acute, some retrorse and curving back slightly. Fixed finger with 40 teeth (♀), 38 (♂). Four to five small, closely spaced teeth just behind venom apparatus, followed by large accessory tooth (Fig. 132). Further teeth arranged in two rows. Movable finger with 27 teeth (♀), 28 (♂). First two teeth behind venom apparatus small, followed by large teeth that decrease in size into small points, basal tooth increased in size into an acute, wide, backwards-facing point.
Chelicera: Hand with five setae located as in Fig. 1. Fixed cheliceral finger with five teeth arranged as in Fig. 129. Female galea complex with nine rami (Fig. 130). Male galea simple and without rami (Fig. 131). Rallum composed of single blade in both sexes. Serrula exterior with 17 lamella (♀) or 16 (♂). Lamina exterior present in both sexes.
Coxae and legs: Pedipalp coxae same colour as carapace, with distinct shoulder. Legs I–IV grey in females, either cream or dark brown in males. Leg coxae yellow to light brown in females, cream or red-brown in males. Leg morphology standard and diplotarsate with regard to both leg I (Fig. 135) and leg IV (Fig. 134).
Measurements: body length ♀ 2.34 ♂ 2.23; carapace ♀ 0.89 x 1.01 (0.88) ♂ 0.90 x 0.86 (1.04). Palp femur ♀ 1.08 x 0.28 (3.85) ♂ 0.99 x 0.26 (3.80), patella ♀ 0.89 x 0.27 (3.29) ♂ 0.83 x 0.26 (3.19), hand ♀ 0.85 x 0.43 ♂ 0.79 x 0.39, chela ♀ 1.62 (3.76) ♂ 1.48 (3.79), movable finger ♀ 0.77 ♂ 0.69. Leg I femur ♀ 0.40 x 0.16 ♂ 0.39 x 0.15, patella ♀ 0.26 x 0.14 ♂ 0.26 x 0.14, tibia ♀ 0.34 x 0.10 ♂ 0.31 x 0.11, metatarsus ♀ 0.24 x 0.08 ♂ 0.27 x 0.08, tarsus ♀ 0.20 x 0.06 ♂ 0.19 x 0.07. Leg IV femur ♀ 0.19 x 0.13 ♂ 0.20 x 0.13, patella ♀ 0.69 x 0.22 ♂ 0.65x 0.22, tibia ♀ 0.55 x 0.13 ♂ 0.53 x 0.13, metatarsus ♀ 0.31 x 0.09 ♂ 0.30 x 0.09, tarsus ♀ 0.28 x 0.07 ♂ 0.22 x 0.07.
Remarks
The second largest geogarypid in South Africa, after G. tectomaculatus sp. nov. The concave depression on the chela was of particular interest to Beier (1947) and was one of the species’ most characteristic features. When examining the type material it was noted that Beier (1947) mentioned four adult specimens in the syntype series, though there were only two specimens in the vial, one female and one male, which are here designated as the lectotype and paralectotype, respectively. The female specimen was in good condition, though severely bleached; the male specimen on the other hand was severely
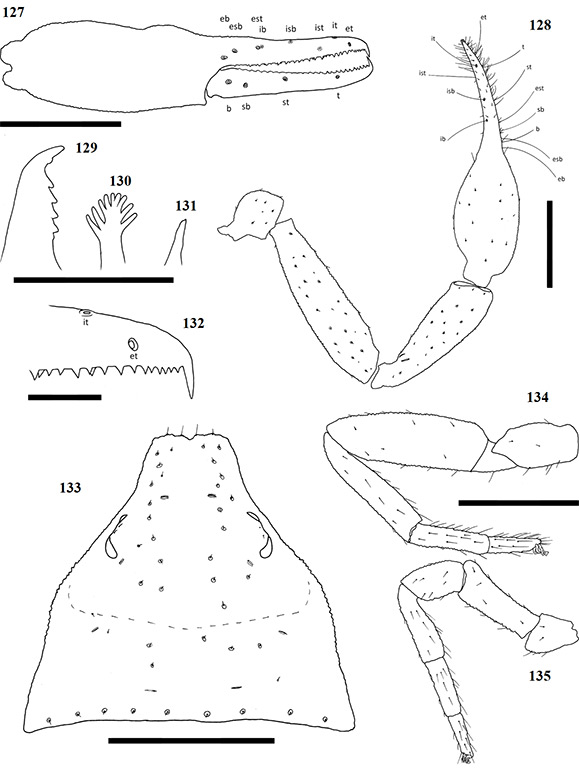
Figures 127–135. Afrogarypus robustus (Beier, 1947) comb. nov. 127–130, 132–135. Female; 131. Male. 127. Right chela, retrolateral view; 128. Right pedipalp, dorsal view; 129. Chelicera movable finger, dorsal view; 130, 131. Galea; 132. Right chela fixed finger, anterior teeth arrangement; 133. Carapace, dorsal view; 134. Right leg IV, prolateral view; 135. Right leg I, prolateral view. Scale bars: Figs 127, 128, 133-135: 0.50 mm; Figs 129-132: 0.10 mm.
damaged. The paratype was in good condition, but also bleached. As with A. purcelli comb. nov., A. minutus comb. nov. and A. triangularis comb. nov., this species is transferred to Afrogarypus due to the presence of a concave dorsal depression on the chelal hand and its grouping within the Afrogarypus clade in the molecular phylogeny.
Additional material examined
SOUTH AFRICA: Western Cape: 1 immature, Cape Town, Kirstenbosch National Botanical Gardens, 33o 59’S, 18o 26’E, 136 m a.s.l., Cultured gardens, leg. J. Pryke, 2.VIII.2005 (WAM T85960); 1♀, Same locality, Skeleton Gorge Trail, 33o 59’S, 18o 25’E, 359 m a.s.l., Afromontane Forest, Leaf litter sifting, leg. J.A. Neethling, 11.XII.2012 (NMBA P00184); 2♀, 3♂, Same data as previous (lactic acid clearing and phylogenetic analysis); 1 immature, Cape Town, Table Mountain, 33o 58’S, 18o 26’E, 360 m a.s.l., Disturbed Fynbos, leg. J. Pryke, 18.I.2007 (WAM T85958); 1♀, Same locality, Natwal Forest South, 34o 00’S, 18o 23’E, leg. J. Pryke, 11.I.2006 (WAM T85954); 1♀, Same locality, Natwal Forest South, 34o 00’S, 18o 25’E, 322 m a.s.l., Exotic forest, leg. J. Pryke, 3.VII.2006 (WAM T85955); 1♀, 1♂, Same locality, Natwal Forest South, 34o 00’S, 18o 23’E, leg. J. Pryke, 13.X.2006 (WAM T85956).
Additional material not examined
SOUTH AFRICA: Western Cape: 1♂, Cape Town, Table Mountain, Wynberg Hollow Gorge [33o 58’S, 18o 25’E], 730 m a.s.l., 18.XII.1950 (examined by Beier 1955).
Afrogarypus subimpressus (Beier, 1955)
Figs 136–148
Geogarypus (Afrogarypus) subimpressus Beier, 1955: 302, fig. 21 right, 22 (Holotype ♂,
South Africa, Western Cape, Simon’s Town, Cape Point Nature Reserve [34o 21’S, 18o 28’E], unknown collector, 10.XII.1950, deposited in NMZA, not examined; Paratypes 2♂, 2 immatures, Same data as previous, deposited in NMZA, not examined).
Geogarypus subimpressus Beier, 1955: Beier, 1964: 60.
Afrogarypus subimpressus (Beier, 1955): Harvey, 1986: 758; Harvey, 1991: 251; Dippenaar- Schoeman & Harvey, 2000: 93.
Diagnosis
Small species. Females can reach a total length (cucullus to posterior abdominal margin) of 1.63 mm, with males reaching 1.46 mm. Carapace dark brown, with slightly lighter posterior in both sexes. All pedipalpal segments same colour as carapace. Well developed, broad sulcus present on dorsal surface of chelal hand. Abdominal tergites of both sexes uniformly brown, with very faded dark patches. Sternites of females weakly sclerotised anteriorly, becoming more strongly sclerotised posteriorly, male sternites well sclerotised throughout. Pedipalp coxae same colour as carapace and possessing a distinct shoulder, legs I–IV pale grey, with the remaining coxae pale brown in colour.
Somewhat resembling A. castigatus sp. nov. in terms of its compact build and colouration, A. subimpressus is easily distinguished by the normal 8/4 trichobothria ratio, diplotarsate front legs and the presence of a well developed sulcus on the dorsal surface of its chela. The dorsal sulcus can be compared with that found in A. megamolaris sp. nov., in that it is broader and more shallow that those found in A. excelsus stat. nov. and A. impressus. Afrogarypus subimpressus can be distinguished from A. megamolaris sp. nov. with regard to its carapace ratio, which is lower than 1 (thus the carapace is broader than long), as well as possessing a prolateral chelal hand surface that is strongly convex, while that of A. megamolaris sp. nov. is relatively straight.
Description
Carapace: Strongly sub-triangular, narrow furrow posterior to the eyes (Fig. 146). Uniformly dark brown with slightly lighter posterior margin in both sexes. Uniformly granular, heavily constricted anteriorly into cucullus, constriction beginning at the medial furrow. Two pairs of corneate eyes situated on ocular tubercles, located about one-third away from the anterior edge. Four prominent setae located on anterior edge, multiple setae located on posterior margin. Numerous small setae present on carapace.
Abdomen: Tergites uniformly brown, indistinct dark patches present in both sexes, granular. Tergites I and II with a large, median, dark brown spot on each. Tergite III without dark patches. Tergites IV–X with paired, indistinct dark patches situated laterally of the midline. Tergites XI and XII without patches. Sternites dark brown in both sexes. Weakly sclerotised anteriorly in females, uniformly sclerotised in males. Pleural membrane wrinkled-plicate, pale.
Pedipalp: Trochanter rounded in shape, uniformly dark brown, granular, small setae scattered over entire surface, except the stem. Apophysis located ventrally. Pedipalp femur (Fig. 141) dark brown, granular. Stem very narrow, widening quickly to form the base, widens only slightly distally before constricting again at the end. Pedipalp patella (Fig. 141) dark brown and granular. Narrow, sharply angled at base, widening quickly into cone. Lyriform fissures visible just anterior to the base, located on bulge, dorso-prolaterally on patella.
Chela: (Figs 136-141) Uniformly dark brown, granular from stem to base of fingers. Both fingers smooth, curving very slightly to interior. Well developed, broad, shallow sulcus present on dorsal surface of chelal hand. Chela hand strongly convex on prolateral edge, retrolateral edge mostly straight. Fixed chela finger with eight trichobothria, movable chela finger with four. Fingers just shorter than hand (with pedicel), venom apparatus present on both.
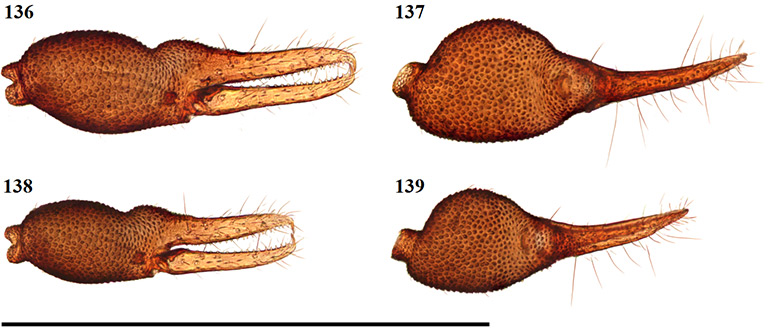
Figures 136–139. Digital microscope photographs of Afrogarypus subimpressus (Beier, 1955) right chela: Female (136, 137) and Male (138, 139). 136, 138. Retrolateral view; 137, 139. Dorsal view. Scale bar: 1.00 mm.
Trichobothria eb, esb and est located retrolaterally in diagonal line at base of fixed chela finger. Trichobothria eb and esb situated about one trichobothrial width from each other, est situated two trichobothrial widths from esb. Trichobothria ib situated prolaterally, just anterior of esb; isb located prolaterally, just anterior of est. Trichobothria ist situated prolaterally and just over one-third away from anterior edge of fixed finger; it located one-quarter finger length from anterior edge, situated dorsally; et located retrolaterally, just anterior of it. Trichobothria b, sb and st located in triangular pattern just anterior of finger base, situated retrolaterally on finger, b situated in line vertically with eb. Trichobothria t located about one-quarter from anterior edge of movable finger, halfway between ist and et. Trichobothria divided in proximal and distal group, proximal group consisting of eb, esb, ib, esb, isb, b, sb and st, distal group containing ist, it, et and t. Chelal teeth acute, slightly retrorse, beginning large anteriorly and gradually decreasing in size to base of finger. Fixed finger with 27 teeth (♀), 24 (♂). First two teeth just before the venom apparatus grouped close together, small (Fig. 142). Further teeth slightly to distinctly retrorse, acute, arranged roughly in two rows. Movable finger with 18 teeth (♀), 17 (♂). First two teeth following the venom apparatus situated on raised ridge. Rest of teeth increase sharply in size posteriorly, distinctly retrorse, additional teeth gradually decreasing in size, spaced further apart.
Chelicera: Hand with five setae situated as in Fig. 1. Fixed cheliceral finger with five teeth arranged as in Fig. 143. Female galea complex, with eight rami in a fan shape (Fig. 144). Male galea simple, no rami (Fig. 145). Rallum composed of single blade in both sexes. Serrula exterior with 15 lamella (♀) or 12 (♂). Lamina exterior present in both sexes.
Coxae and legs: Pedipalp coxae dark brown, possessing distinct shoulder. Legs I–IV light grey. Joint between metatarsus and tarsus of legs I and II almost completely fused, immovable, slight fusion line visible (Fig. 147). Leg pairs III and IV diplotarsate (Fig. 148).
Measurements: body length ♀ 1.63 ♂ 1.46; carapace ♀ 0.51 x 0.57 (0.89) ♂ 0.44 x 0.47 (0.93). Palp femur ♀ 0.52 x 0.16 (3.25) ♂ 0.45 x 0.14 (3.21), patella ♀ 0.40 x 0.15 (2.66) ♂ 0.35 x 0.14 (2.50), hand ♀ 0.39 x 0.26 ♂ 0.31 x 0.22, chela ♀ 0.75 (2.88) ♂ 0.63 (2.86), movable finger ♀ 0.35 ♂ 0.29. Leg I femur ♀ 0.24 x 0.08 ♂ 0.19 x 0.08, patella ♀ 0.12 x 0.08 ♂ 0.11 x 0.08, tibia ♀ 0.16 x 0.06 ♂ 0.13 x 0.06, metatarsus ♀ 0.12 x 0.05 ♂ 0.11 x 0.04, tarsus ♀ 0.10 x 0.03 ♂ 0.10 x 0.03. Leg IV femur ♀ 0.11 x 0.07 ♂ 0.10 x 0.06, patella ♀ 0.32 x 0.13 ♂ 0.28 x 0.11, tibia ♀ 0.27 x 0.08 ♂ 0.26 x 0.07, metatarsus ♀ 0.14 x 0.06 ♂ 0.13 x 0.05, tarsus ♀ 0.13 x 0.04 ♂ 0.13 x 0.03.
Remarks
When Beier (1955) described this species it was known only from the Cape Peninsula, although it has since been shown to have a much broader distribution. Due to the presence of a well developed dorsal sulcus, then only known from A. impressus, and owing to its diminutive size, Beier opted to name the species A. subimpressus, combining both the size and relationship to A. impressus in one name. The species has been in Afrogarypus since its
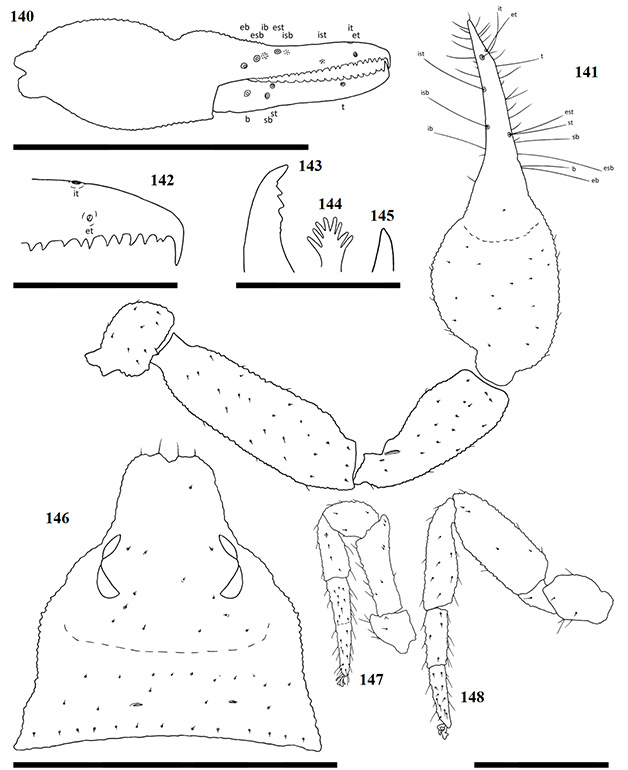
Figures 140–148. Afrogarypus subimpressus (Beier, 1955). 140–144, 146–148. Female; 145. Male. 140. Right chela, retrolateral view; 141. Right pedipalp, dorsal view; 142. Right chela fixed finger, anterior teeth arrangement; 143. Chelicera movable finger, dorsal view; 144, 145. Galea; 146. Carapace, dorsal view; 147. Right leg I, prolateral view; 148. Right leg IV, prolateral view. Scale bars: Figs 140, 141, 146-148: 0.50 mm; Figs 142-145: 0.10 mm.
description, although Afrogarypus was then still a subgenus of Geogarypus. It was consequently retained in Afrogarypus in the revision by Harvey (1986) and given its current status as A. subimpressus. Although the species was collected from the type locality of A. purcelli comb. nov. (Beaufort West), it can clearly be separated from the latter species by its well defined dorsal sulcus, as well as by lacking the indentation on the prolateral surface of the chela.
Additional material examined
SOUTH AFRICA: Free State: 2♀, Luckhof District, Bankfontein Farm, 30o 04’S, 24o 53’E, 1186 m a.s.l., leg. UFS Entomology Students, 2.IV.2015 (NMBA P00237); 1♂, Bethulie, Tussen-die-Riviere Nature Reserve, 30o 29’S, 26o 07’E, 1288 m a.s.l., leg. L.N. Lotz & C.R. Haddad, 16.X.2008 (NMBA P00135). Northern Cape: 1♂, Hanover, 31o 15’S, 24o 24’E, leg. J.M. Waldock, 17.IV.2001 (WAM T67018). Western Cape: 1♀, 3♂, Beaufort West, Karoo National Park, Klipspringer Pass, 32o 19’S, 22o 27’E, 1126 m a.s.l., leg. J.A. Neethling, 14.XI.2013 (NMBA P00175); 2♂, 1 immature, Cape Town, Table Mountain, 33o 58’S, 18o 23’E, leg. J. Pryke, 17.X.2005 (WAM T85951, T85952 & T85953); 1 Deutonymph, Knysna, Brenton-on-Sea [34o 03’S, 22o 59’E], leg. H.G. Robertson, 23.XI.1998 (SAMC ENW-COO5851); 2♀, 3♂, Simon’s Town, Cape Point Nature Reserve, Cape Point, 34o 21’S, 18o 29’E, 118 m a.s.l., leg. J.A. Neethling, 26.IV.2013 (NMBA P00174); 3♀, 3♂, Same data as previous (morphological dissection and phylogenetic analysis); 1♀, 2♂, Same locality, Olifantsbos, 34o 15’S, 18o 22’E, 9 m a.s.l., leg. J.A. Neethling, 12.XII.2012 (NMBA P00176); 1♀, 2 Deutonymphs, Same locality, Olifantsbos, 34o 15’S, 18o 23’E, leg. H.G. Robertson, X.1998 (SAMC ENW-COO 5432); 3♀, 3♂, 2 Deutonymphs, Same locality, Smitswinkelvlakte [34o 16’S, 18o 25’E], leg. H.G. Robertson, X.1998 (SAMC ENW-COO 5404, 5407, 5408 & 5436).
Additional material not examined
SOUTH AFRICA: Unknown: 2♀, 1♂, No collection information (examined by Beier 1964, deposited in NMZA).
Afrogarypus triangularis (Ellingsen, 1912) comb. nov.
Figs 149–162
Garypus minutus var. triangularis Ellingsen, 1912: 110 (Holotype Unknown gender, South
Africa, Eastern Cape, King William’s Town, Izeli [32o 48’S, 27o 23’E], No further data, not examined; Paratypes 1 specimen, South Africa, Eastern Cape, Frankfort, Frankfort Hill [32o 43’S 27o 27’E], No further data, not examined; 1 specimen, South Africa, Eastern Cape, Butterworth, Blythswood, Bushman’s Rock [32o13’S, 27o58’E], No further data, not examined; 2 specimens, South Africa, Free State, Bloemfontein, Naval Hill [29o 06’S, 26o 14’E], No further data, not examined).
Geogarypus (Geogarypus) minutus var. triangularis (Ellingsen, 1912): Beier, 1932: 23.
Geogarypus triangularis (Ellingsen, 1912): Roewer, 1937: 269; Beier, 1964: 59, fig. 20;
Harvey, 1986: 760; Harvey, 1991: 260; Dippenaar-Schoeman & Harvey, 2000: 93 [comb. nov.].
Diagnosis
Medium sized species. Females can reach a total length (cucullus to posterior abdominal margin) of 2.10 mm, with males reaching up to 2.08 mm. Carapace uniform dark brown in females, brown with medial furrow and posterior margin light brown in males. All pedipalp segments brown in colour. Concave depression present on dorsal surface of chela. Abdominal tergites mostly brown with dark patches, abdominal sternites light brown and weakly sclerotised (♀), light brown and well sclerotised throughout (♂). Males with distinct lateral sclerites between tergites and sternites. Pedipalp coxae brown in colour with distinct shoulder, legs I–IV as well as remaining coxae tan to pale yellow. Both sexes are quite variable in size, although males tend to be smaller than the females from the same location.
Smaller A. triangularis comb. nov. specimens closely resemble A. minutus comb. nov. with regard to size and colouration, but can be distinguished by the triangular shape of the chela hand (only somewhat convex or rectangular in A. minutus comb. nov.). Females can furthermore be identified by the differences in galea structure, while adult males can easily be differentiated by the presence of lateral sclerites on the abdomen. Together with A. castigatus sp. nov., they are the only known South African geogarypids that possess lateral abdominal sclerites, a trait they share with members of the Feaellidae. In both A. triangularis comb. nov. and A. castigatus sp. nov. only mature males have these sclerites.
Description
Carapace: Strongly sub-triangular, narrow furrow posterior to the eyes (Fig. 157). Uniformly dark brown in females, overall brown with light brown medial furrow and posterior margin in males. Uniformly granular, heavily constricted anteriorly into cucullus, constriction beginning at the medial furrow. Two pairs of corneate eyes situated on ocular tubercles, located about one-third away from anterior edge. Four prominent setae located on anterior edge, row of 12 setae, seated within rims, located on posterior margin. Numerous small setae present on carapace.
Abdomen: Abdominal tergites granular, brown with dark patches in both sexes. Tergites I and II with large medial dark spot each. Tergite III without dark patches. Tergites IV–X with paired, indistinct dark patches just lateral of midline. Tergites XI and XII without dark patches. Sternites light brown, weakly sclerotised (♀). Darker brown, well sclerotised throughout (♂). Pleural membrane wrinkled-plicate, males with 17 lateral sclerites (Fig. 158) on each side of abdomen, arranged as follows: one sclerite beneath tergite III, two beneath tergite IV, three beneath tergite V, two beneath tergite VI, three beneath both tergites VII and VIII, two beneath tergite IX , one beneath tergite X.
Pedipalp: Trochanter rounded in shape, granular, uniformly dark brown. Small setae scattered over entire surface, except the stem. Apophysis located ventrally. Pedipalp femur (Fig. 154) dark brown and granular. Narrower at base, widening evenly before constricting again at end. Pedipalp patella (Fig. 154) dark brown and granular. Narrow, sharply angled at base, widening sharply into cone shape. Lyriform fissures visible just anterior of base, located on bulge, dorso-prolaterally on patella.
Chela: (Figs 149-154) Uniformly brown, granular from stem to base of fingers. Both fingers smooth. Concave depression present on dorsal surface of chela, resulting in bulge at base of fixed finger. Chela hand strongly convex on prolateral edge, blunt constriction just posterior to base of fingers giving chela somewhat triangular appearance. Fixed chela finger with eight trichobothria, movable chela finger with four, both fingers narrow, curving slightly to interior. Fingers just shorter than hand (with pedicel). Venom apparatus present on both fingers. Trichobothria eb, esb and est located retrolaterally in diagonal line at base of fixed chela finger, situated about one trichobothrial width from each other. Trichobothria ib situated dorso-prolaterally, nearly exactly opposite of est; isb located dorso-prolaterally, one trichobothrial width forward of ib. Trichobothria ist situated prolaterally, one-third away from anterior edge of fixed finger; it located one-quarter of finger length from anterior edge, situated dorsally; et located retrolaterally and distally from it, nearly equidistant from it as ist from it. Trichobothria b, sb and st located in triangular pattern just anterior to finger base, situated retrolaterally on finger, b situated in line vertically with eb. Trichobothria t located about. One-third from the anterior edge of movable finger, in line vertically with ist. Trichobothria can be divided into two groups, with proximal group consisting of eb, esb, ib, esb, isb, b, sb and st, distal group containing ist, it, et and t. Chelal teeth acute, mostly retrorse, some curving back. Fixed finger with 22 teeth (♀), 19 (♂). First tooth just before venom apparatus not retrorse, almost pointing anteriorly (Fig. 159). Further teeth slightly to distinctly retrorse and acute, arranged roughly in two rows. Movable finger with 16 teeth (♀), 18 (♂). First five teeth following the venom apparatus in females gradually increase in size, rest of teeth gradually decrease in size, spacing further apart. In males, first 11 teeth roughly the same size, remaining seven teeth being reduced to acute points.
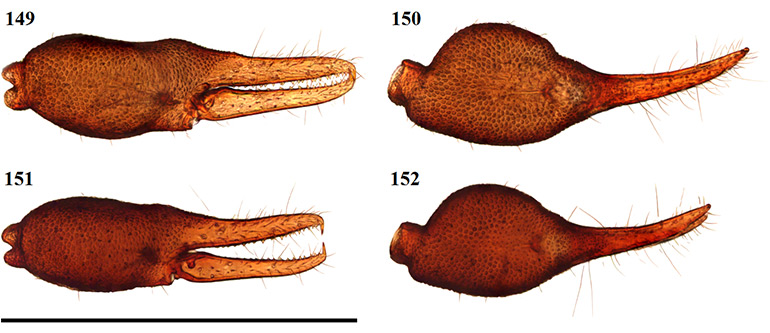
Figures 149–152. Digital microscope photographs of Afrogarypus triangularis (Ellingsen, 1912) comb. nov. right chela: Female (149, 150) and Male (151, 152). 149, 151. Retrolateral view; 150, 152. Dorsal view. Scale bar: 1.00 mm.
Chelicera: Hand with five setae as in Fig. 1. Fixed cheliceral finger with five teeth arranged as in Fig. 160. Female galea complex with nine rami, basal pair situated small distance from remaining group as in Fig. 161. Male galea simple, no rami (Fig. 162). Rallum composed of single blade in both sexes. Serrula exterior with 16 lamella (♀) or 13 (♂). Lamina exterior present in both sexes.
Coxae and legs: Pedipalp coxae brown in colour, shoulder distinct. Multiple long setae present along inner medial ridges. Coxae of legs I–IV, as well as all leg segments, tan. Trochanters of legs I and II compact and small, larger and more elongate on legs III and IV. Femora of legs I and II longer than patellae and tibiae respectively (Fig. 155). Metatarsi and tarsi not fused, joint movable. Tarsal claws simple, curved, arolium much longer than claws, not divided. Femur-patella joint of leg IV immovable, tibia mostly straight (Fig. 156), metatarsi and tarsi not fused, joint movable. Claws and arolium similar to those of legs I–III.
Measurements: body length ♀ 2.10 ♂ 2.08; carapace ♀ 0.68 x 0.67 (1.01) ♂ 0.64 x 0.63 (1.01). Palp femur ♀ 0.65 x 0.19 (3.42) ♂ 0.59 x 0.19 (3.10), patella ♀ 0.48 x 0.18 (2.66) ♂ 0.44x 0.19 (2.31), hand ♀ 0.53 x 0.34 ♂ 0.47 x 0.32, chela ♀ 1.01 (2.97) ♂ 0.89 (2.78), movable finger ♀ 0.44 ♂ 0.42. Leg I femur ♀ 0.27 x 0.12 ♂ 0.24 x 0.11, patella ♀ 0.16 x 0.11 ♂ 0.14 x 0.10, tibia ♀ 0.21 x 0.09 ♂ 0.18 x 0.08, metatarsus ♀ 0.13 x 0.06 ♂ 0.12 x 0.06, tarsus ♀ 0.15 x 0.04 ♂ 0.14 x 0.04. Leg IV femur ♀ 0.14 x 0.12 ♂ 0.13 x 0.09, patella ♀ 0.41 x 0.17 ♂ 0.39 x 0.15, tibia ♀ 0.39 x 0.11 ♂ 0.38 x 0.11, metatarsus ♀ 0.19 x 0.07 ♂ 0.18 x 0.07, tarsus ♀ 0.17 x 0.06 ♂ 0.14 x 0.06.
Remarks
This species was originally regarded as a variant of A. minutus comb. nov. by Ellingsen (1912) due to their similarities, but he separated A. triangularis comb. nov. due to its distinctive triangular chela. The species then continued to hold its subspecies status until it was properly re–described by Beier (1964), who elevated it to full species status as G. triangularis. As the species possesses a concave dorsal depression on the chelal hand and groups within the Afrogarypus clade in the molecular phylogeny, similar to A. purcelli comb. nov., A. minutus comb. nov and A. robustus comb. nov., it is transferred to Afrogarypus.
The original variant was identified from a group of nine immature specimens that Ellingsen received from Reverend Robert Godfrey’s collection, originally housed at the Iziko South African Museum. He unfortunately never mentions the gender or the depository numbers of the types, providing only the sampling locations of the nine specimens. After extensive searching in the collections housed in the SAMC, neither the type specimens for A. triangularis comb. nov.,
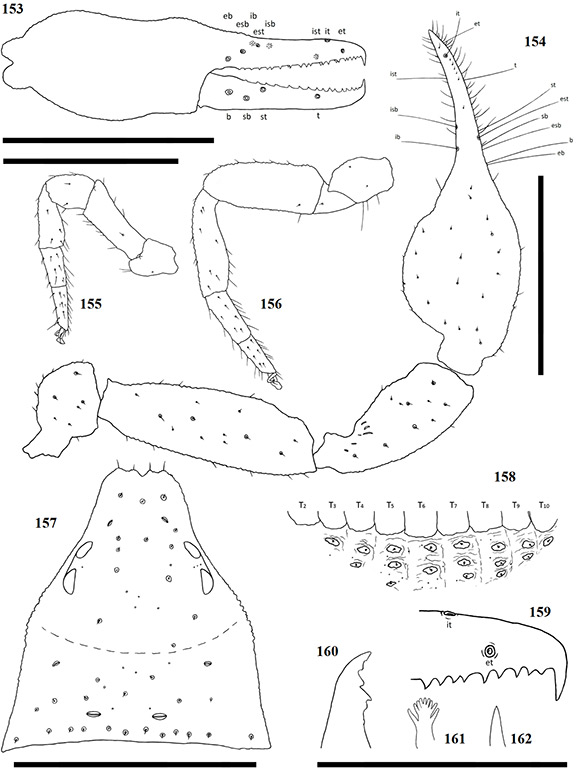
Figures 153–162. Afrogarypus triangularis (Ellingsen, 1912) comb. nov. 153–157, 159–161. Female; 158, 162. Male. 153. Right chela, retrolateral view; 154. Right pedipalp, dorsal view; 155. Leg I, prolateral view; 156. Leg IV, prolateral view; 157. Carapace, dorsal view; 2158. Male abdominal pleural sclerites; 159. Chela fixed finger, anterior teeth arrangement; 160. Chelicera movable finger, dorsal view; 161, 162. Galea. Scale bars: Figs 153-157: 0.50 mm; Figs 159-162: 0.10 mm.
nor those of A. purcelli comb. nov. (also originally housed in this museum), could be located. Even after collaborated searches in prominent museum collections overseas, the type specimens of these two species could not be located. However, adequate material has been collected from the vicinity of the type locality to verify the status and identity of the species. The Bloemfontein, Free State, record is suspected to be a mis-identification, as no specimens of the species could be located on Naval Hill and surrounding areas, even after multiple attempts by the author and Dr. L.N. Lotz of the National Museum Bloemfontein.
Additional material examined
SOUTH AFRICA: Eastern Cape: 1♀, Burgersdorp [30o 59’S 26o 19’E], leg. R.F. Lawrence, X.1944 (NMSA 677); 3♀, 4♂, Cradock, Mountain Zebra National Park, bushveld, 32o 13’S, 25o 28’E, 1240 m a.s.l., leg. J.A. Neethling & C. Luwes, 29.III.2012 (NMBA P00187); 13 Adults, Grahamstown, Grey Dam [33o 19’S 26o 31’E], leg. J. Hewitt, 3.X.1918 (AMG Slide); 2 Adults, Same locality, leg. J. Hewitt, XII.1918 (AMG Slide); 11♀, 8♂, Hogsback, Hogsback Arboretum, invaded forest, 32o 35’S, 26o 56’E, 1247 m a.s.l., leg. J.A. Neethling & C. Luwes, 2.IV.2012 (NMBA P00182); 1♀, Same locality, Pine Plantation, 32o 36’S, 26o 56’E, 1205 m a.s.l., leg. UFS Entomology Students, 30.III.2009 (NMBA P00146); 7♀, 6♂, King William’s Town, Mount Coke State Forest, indigenous forest, 32o 59’S, 27o 28’E, 420 m a.s.l., leg. J.A. Neethling, 30.XI.2013 (NMBA P00183); 2♀, 2♂, Same data as previous (morphological dissection and phylogenetic analysis).
Additional material not examined
SOUTH AFRICA: Eastern Cape: 1♀, Grahamstown, Coldsprings, Southern slope [33o 06’S, 26o 41’E], V.1920 (examined by Beier 1964, deposited in NMZA); 1♀, Port Elizabeth [33o 56’S, 25o 34’E], leg. F. Emden (examined by Beier 1964, deposited in NMZA). Unknown: 7♀, 5♂, No collection information, (examined by Beier 1964, deposited in NMZA).
Geogarypus deceptor sp. nov.
Figs 163–178
Holotype: ♀, SOUTH AFRICA, KwaZulu-Natal, St. Lucia, iSimangaliso Wetland Park, Cape Vidal, 28o 08’S, 32o 33’E, 62 m a.s.l., Coastal forest, Leaf litter sifting, leg. J.A. Neethling & C. Luwes, 12.V.2012 (NMBA P00221). Paratypes: 1♀, 2♂, Same data as holotype (NMBA P00222).
Etymology
The Latin noun deceptor means deceiver and refers specifically to all the examined museum exemplars which either lacked a species level identification, or were misidentified as Geogarypus olivaceus.
Diagnosis
Medium sized species. Both males and females reach a length (cucullus to posterior abdominal margin) of 1.65 mm. Carapace dark brown with a large cream patch on the posterior in both sexes. Pedipalp segments same colour as carapace, with the dorsal surface of the chela hand convex from the stem to the base of the chelal fingers. Abdominal tergites brown to light brown in colour, with dark and cream patches in both sexes, while the abdominal sternites are light brown with distinct dark spots in both sexes. Pedipalp coxae same colour as carapace with distinct shoulder, legs I–IV with corresponding coxae cream to tan in colour in both sexes.
This species closely resembles both G. liomendontus sp. nov. and G. modjadji sp. nov. in both morphology and colouration, but can be distinguished from them by the lack of any fused chelal teeth, as well as possessing an isolated basal tooth close to the base of the movable chelal finger. Although both sexes can reach the same average length, females possess longer appendages than males.
Description
Carapace: Strongly sub-triangular, narrow furrow posterior to the eyes (Fig. 176). Carapace dark brown, large dorsal cream patch behind medial furrow, uniformly granular in both sexes. Heavily constricted anteriorly into cucullus, constriction beginning at medial furrow. Two pairs of corneate eyes situated on ocular tubercles, located about one-third from anterior edge. Four prominent setae located on anterior edge, row of 10 setae located on posterior margin. Numerous small setae present on carapace.
Abdomen: Abdominal tergites brown to light brown with both dark and cream patches on most plates. Tergites I and II with large median dark spot each, flanked by cream patches. Tergite III without dark patches, almost entirely cream. Tergites IV–X with cream posterior margins together with paired dark patches just lateral of the midline. Tergites XI and XII lack dark patches. Setae of posterior setal rows of tergites situated on small pale plates with dark edges, making plates distinctly visible. Sternites light brown, well sclerotised in both sexes, paired dark spots located laterally of midline. Pleural membrane wrinkled-plicate, cream.
Pedipalp: Trochanter rounded anteriorly, granular, same dark colouration as carapace. Small setae located on dorsal surface, ventral apophysis present. Pedipalp femur (Fig. 170) and patella same colour as trochanter, granular. Femur narrow at base, widening evenly until just posterior of anterior margin, where there is a sudden constriction. Pedipalp patella (Fig. 170) narrow, slightly angled at base, widening evenly into cone. Lyriform fissures visible just anterior of base, located on bulge, dorso-prolaterally on patella.
Chela: (Figs 163–166, 169, 170) Chela palm same colour as rest of the pedipalp, fingers lighter in both sexes, granular from stem up to base of fingers. Both fingers smooth, slightly shorter than hand (with pedicel). Dorsal surface of chelal hand uniformly convex in shape from stem to base of chelal fingers. Both pro- and retrolateral surfaces slightly convex, prolateral surface more so. Fixed chela finger with eight trichobothria, movable chela finger with four. Both fingers long and narrow, curving to interior. Venom apparatus present on both fingers.
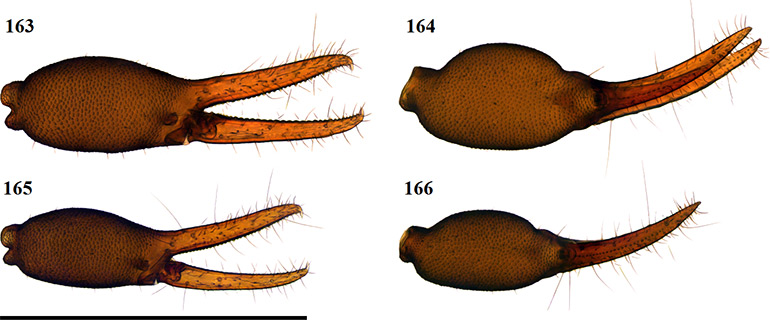
Figures 163–166. Digital microscope photographs of Geogarypus deceptor sp. nov. right chela: Female (163, 164) and Male (165, 166). 163, 165. Retrolateral view; 164, 166. Dorsal view. Scale bar: 1.00 mm.
Trichobothria eb and esb one trichobothrial width from each other, located retrolaterally at base of fixed finger, ib located dorso-prolaterally, two trichobothrial widths distal of esb. Trichobothria est located one-quarter finger length on retrolateral surface, with isb situated distally, located two-fifths finger length from base of fixed finger on dorsal surface. Trichobothria ist located one-third finger length from anterior finger edge, roughly in middle of the prolateral surface, it located on dorsal surface just over one-quarter finger length from anterior finger edge; et situated anteriorly on retrolateral surface. Trichobothria b and eb on movable finger situated somewhat forward of finger base, b ventrally in line with esb. Trichobothria st is positioned in line ventrally with isb, t located roughly one-quarter finger length from anterior edge of movable finger. Chelal teeth acute and retrorse. Fixed finger with 32 teeth (♀), 37 (♂). All teeth are situated in one row, several large teeth located equally along finger length (Fig. 172). Movable finger with 29 teeth (♀), 30 (♂). Basal teeth on female movable finger increase in size into acute points, both sexes possessing an isolated tooth at base of movable finger (Figs 167, 168, 171).

Figures 167–168. Digital microscope photographs of Geogarypus deceptor sp. nov., arrows indicating isolated basal chelal tooth of female (167) and male (168).
Chelicera: Hand with five setae located as in Fig. 1. Fixed cheliceral finger with five teeth arranged as in Fig. 173. Female galea complex with nine rami (Fig. 174). Male galea simple, with two spinules (Fig. 175). Rallum composed of single blade in both sexes. Serrula exterior with 17 lamella in both sexes. Lamina exterior present in both sexes.
Coxae and legs: Pedipalp coxae same colour as carapace, with distinct shoulder, legs I–IV with corresponding coxae cream to tan in both sexes. All legs diplotarsate (Figs 177 & 178).
Measurements: body length ♀ 1.65 ♂ 1.65; carapace ♀ 0.73 x 0.75 (0.97) ♂ 0.62 x 0.58 (1.06). Palp femur ♀ 0.75 x 0.23 (3.26) ♂ 0.69 x 0.22 (3.15), patella ♀ 0.59 x 0.22 (2.68) ♂ 0.54 x 0.21 (2.58), hand ♀ 0.65 x 0.38 ♂ 0.54 x 0.29, chela ♀ 1.29 (3.39) ♂ 1.10 (3.79), movable finger ♀ 0.62 ♂ 0.55. Leg I femur ♀ 0.33 x 0.12 ♂ 0.30 x 0.11, patella ♀ 0.20 x 0.11 ♂ 0.17 x 0.11, tibia ♀ 0.26 x 0.08 ♂ 0.22 x 0.08, metatarsus ♀ 0.18 x 0.06 ♂ 0.17 x 0.06, tarsus ♀ 0.17 x 0.04 ♂ 0.16 x 0.05. Leg IV femur ♀ 0.15 x 0.11 ♂ 0.14 x 0.11, patella ♀ 0.52 x 0.18 ♂ 0.43 x 0.17, tibia ♀ 0.42 x 0.11 ♂ 0.38 x 0.11, metatarsus ♀ 0.24 x 0.07 ♂ 0.21 x 0.08, tarsus ♀ 0.22 x 0.06 ♂ 0.18 x 0.05.
Remarks
This is the only South African species that possesses an isolated basal tooth on the movable chela finger. Not many museum exemplars were found; all were either unidentified or mis-identified as G. olivaceus.
Additional material examined
SOUTH AFRICA: KwaZulu-Natal: 2♀, 1♂, Jozini, Jozini Town Outskirts, 27o 29’S, 32o 01’E, 454 m a.s.l., Indigenous bush patch, Leaf litter sifting, leg. J.A. Neethling & C. Luwes, 19.V.2012 (NMBA P00218); 3♀, 2♂, Mbazwana, iSimangaliso Wetland Park, Sodwana Bay National Park, 27o 32’S, 32o 41’E, 11 m a.s.l., Coastal bush, Leaf litter sifting, leg. J.A. Neethling & C. Luwes, 18.V.2012 (NMBA P00217); 2♀, 1♂, Ndumo Game Reserve, 26o 55’S, 32o 16’E, 120 m a.s.l., Under rocks, leg. C.R. Haddad, 7.I.2007 (WAM T67148, T79040, T79039); 1♀, Same locality, Crocodile Farm, 26o 54’S, 32o 19’E, 57 m a.s.l., leg. C.R. Haddad, 7.II.2005 (WAM T79038); 1♀, Ithala Game Reserve, Ntshodwe Camp, 27o 33’S, 31o 17’E, 1170 m a.s.l., leg. C.R. Haddad & Z. Mbo, 29.I.2014 (NMBA P00196); 3♀, Umhlali, Sheffield Beach [29o 29’S, 31o 15’E], leg. R.F. Lawrence, VII.1958 (NMSA 5163); 2♀, 5♂, 1 Deutonymph, St. Lucia, Dukuduku Forest [28o 22’S, 32o 20’E], leg. R.F. Lawrence & J.Y. Lawrence, XII.1960 (NMSA 7864); 3♀, 5♂, St. Lucia, St. Lucia Coastal Forest, 28o 23’S, 32o 24’E, 20 m a.s.l., Coastal forest, Leaf litter sifting, leg. J.A. Neethling & C. Luwes, 13.V.2012 (morphological dissection and phylogenetic analysis); 2♀, Tembe Elephant Park, 27o 03’S, 32o 27’E, 68 m a.s.l., Closed woodland, leg. C.R. Haddad, 6.I.2002 (WAM T86753). Mpumalanga: 2♀, Kruger National Park, Pretoriuskop Rest Camp, [25o 10’S, 31o 16’E], leg. R.F. Lawrence, XII.1960 (NMSA 16048); 1♀, 1♂, Nelspruit, Lowveld National Botanical Gardens, 25o 27’S, 30o 58’E, 642 m a.s.l., Indigenous forest patch, Leaf litter sifting, leg. J.A. Neethling, 1.X.2012 (NMBA P00214).

Figures 169–178. Geogarypus deceptor sp. nov. 169–174, 176–178. Female; 175. Male. 169. Right chela, retrolateral view; 170. Right pedipalp, dorsal view; 171. Basal teeth arrangement; 172. Anterior teeth arrangement on fixed chelal finger; 173. Chelicera movable finger, dorsal view; 174, 175. Galea; 176. Carapace, dorsal view; 177. Right leg IV, prolateral view; 178. Right leg I, prolateral view. Scale bars: Figs 169, 170, 176-178: 0.50 mm; Figs 172-175: 0.10 mm.
Geogarypus flavus Beier, 1947 stat. nov.
Figs 179-192
Geogarypus (Geogarypus) flavus Beier, 1947: 318, fig. 24 (Holotype ♀, South Africa,
Western Cape, Plettenberg Bay, Keurbooms River [33o 59’S, 23o 24’E], leg. K.H.B. [SAMC B6922], examined); Beier, 1958: 170.
Geogarypus olivaceus (Tullgren, 1907): Beier, 1964: 59 (synonomised).
Diagnosis
Large species. Females can reach a length (cucullus to posterior abdominal margin) of 2.92 mm, with males reaching 2.74 mm. Carapace dark brown in both sexes, with a lighter coloured band around the medial furrow and posterior edge of the carapace. Pedipalp segments same colour as carapace, with the dorsal surface of the chela hand convex from the stem to the base of the chelal fingers. Abdominal tergites mostly medium brown to dark brown, with dark and cream patches, while the abdominal sternites are medium brown to light brown with dark spots, and well sclerotised in both sexes. Pedipalp coxae same colour as carapace with distinct shoulder, legs I–IV with corresponding coxae yellow to light brown in both sexes.
Longer and more narrow, but less bulky than A. robustus comb. nov., G. flavus stat. nov. is one of the largest geogarypid species in South Africa. Large specimens closely resemble G. tectomaculatus sp. nov. in general size, morphology and colouration, while smaller specimens resemble G. olivaceus. In both instances, G. flavus stat. nov. can be distinguished by having small, separated basal chelal teeth, while the basal teeth of G. olivaceus and G. tectomaculatus sp. nov. are larger and closely spaced.
Description
Carapace: Strongly sub-triangular, longer than broad in both sexes, with narrow furrow posterior to the eyes (Fig. 190). Carapace dark brown, uniformly granular in both sexes, lighter coloured band around medial furrow and posterior edge. Heavily constricted anteriorly into cucullus, constriction beginning at medial furrow. Two pairs of corneate eyes situated on ocular tubercles, located about one-third away from anterior edge. Four prominent setae located on anterior edge, row of 10 setae located on posterior margin. Numerous small setae present on carapace.
Abdomen: Abdominal tergites medium brown to dark brown, both dark and cream coloured patches on most plates. Tergites I and II with large median dark spot each, flanked by cream patches. Tergite III without dark patches, with median cream band extending almost entire length of tergum. Tergites IV–X with medial cream patch and paired dark patches just lateral of midline, flanked laterally by cream patches. Tergites XI and XII lack dark patches. Setae of posterior setal rows of tergites, predominantly in females, situated on large pale plates with dark edges, making these plates distinctly visible. Sternites well sclerotised in both sexes, paired dark spots located laterally of cream midline. Pleural membrane wrinkled-plicate, cream.
Pedipalp: Trochanter rounded anteriorly, granular, same colouration as carapace. Small setae located on dorsal and dorso-lateral sides, apophysis located ventrally. Pedipalp femur (Fig. 184) and patella same colour as trochanter, granular. Femur narrow at base, widening evenly until just posterior of anterior margin, where there is a sudden constriction. Pedipalp patella (Fig. 184) narrow, slightly angled at base, widening evenly into a cone. Lyriform fissures visible just anterior to base, located on bulge, dorso-prolaterally on patella.
Chela: (Figs 179-184) Mostly same colour as rest of pedipalp, can be slightly lighter in both sexes. Granular from stem up to base of fingers. Both fingers smooth, just as long as hand (with pedicel). Dorsal surface of chelal hand uniformly convex from stem to base of chelal fingers. Both pro- and retrolateral surfaces slightly convex. Fixed chela finger with eight trichobothria, movable chela finger with four. Both fingers long, narrow, curving to the interior. Venom apparatus present on both fingers. Trichobothria eb and esb situated one trichobothrial width from each other, retrolaterally at base of fixed finger; ib located dorso-prolaterally, three trichobothrial widths distal of esb. Trichobothria est located one-third finger length on retrolateral surface, isb situated distally by three trichobothrial widths on opposite surface. Trichobothria ist located one-third finger length from anterior finger edge, roughly in middle of prolateral surface; it located on dorsal surface, one-fifth finger length from anterior finger edge, et situated just anterior on retrolateral surface. Trichobothria b and eb on movable finger, somewhat forward of finger base, b ventrally in line with esb. Trichobothria st positioned roughly halfway along movable finger, t located roughly one-quarter finger length from anterior edge of movable finger. Chelal teeth acute and retrorse. Fixed finger with 41 teeth (♀), 40 (♂). First three teeth behind venom apparatus raised, forming distinct group (Fig. 189). Formation then followed by remaining fixed chelal teeth, within a single row, only decreasing slightly in size to base (Fig. 188). Movable finger with 30 teeth (♀), 31 (♂). First tooth behind venom apparatus small, followed by larger teeth that decrease in size into small points. In both sexes the first few basal teeth on movable finger are fully formed/are present as wide teeth points.
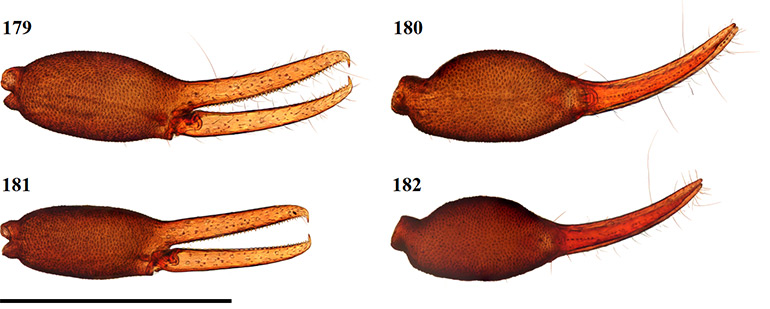
Figures 179–182. Digital microscope photographs of Geogarypus flavus (Beier, 1947) stat. nov. right chela: Female (179, 180) and Male (181, 182). 179, 181. Retrolateral view; 180, 182. Dorsal view. Scale bar: 1.00 mm.
Chelicera: Hand with five setae located as in Fig. 1. Fixed cheliceral finger with six teeth arranged as in Fig. 185. Female galea complex with nine rami (Fig. 186), simple, with two spinules in males (Fig. 187). Rallum composed of single blade in both sexes. Serrula exterior with 19 lamella in both sexes. Lamina exterior present in both sexes.
Coxae and legs: Pedipalp coxae same colour as trochanters, with distinct shoulder. Legs I–IV with corresponding coxae yellow to light brown. All legs diplotarsate (Figs 191 & 192).
Measurements: body length ♀ 2.97 ♂ 2.74; carapace ♀ 0.88 x 0.84 (1.04) ♂ 0.74 x 0.69 (1.07). Palp femur ♀ 0.92 x 0.27 (3.40) ♂ 0.88 x 0.25 (3.52), patella ♀ 0.74 x 0.27 (2.74) ♂ 0.70 x 0.25 (2.80), hand ♀ 0.75 x 0.41 ♂ 0.67 x 0.37, chela ♀ 1.54 (3.75) ♂ 1.36 (3.67), movable finger ♀ 0.77 ♂ 0.69. Leg I femur ♀ 0.38 x 0.16 ♂ 0.35 x 0.13, patella ♀ 0.23 x 0.14 ♂ 0.19 x 0.12, tibia ♀ 0.31 x 0.10 ♂ 0.26 x 0.09, metatarsus ♀ 0.20 x 0.08 ♂ 0.18 x 0.07, tarsus ♀ 0.19 x 0.05 ♂ 0.17 x 0.05. Leg IV femur ♀ 0.18 x 0.13 ♂ 0.19 x 0.13, patella ♀ 0.60 x 0.22 ♂ 0.54 x 0.21, tibia ♀ 0.49 x 0.13 ♂ 0.42 x 0.13, metatarsus ♀ 0.25 x 0.09 ♂ 0.24 x 0.08, tarsus ♀ 0.21 x 0.06 ♂ 0.22 x 0.06.
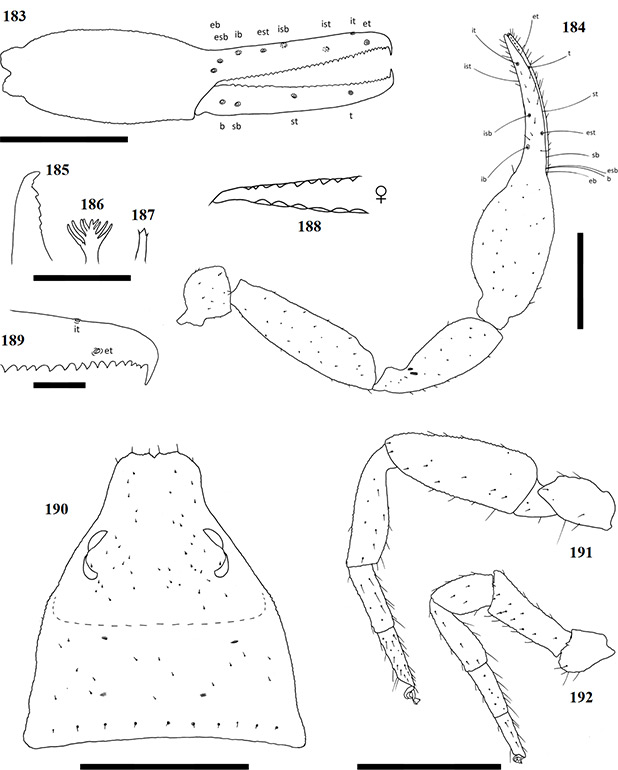
Figures 183–192. Geogarypus flavus (Beier, 1947) stat. nov. 183–186, 188–192. Female; 187. Male. 183. Right chela, retrolateral view; 184. Right pedipalp, dorsal view; 185. Chelicera movable finger, dorsal view; 186, 187. Galea; 188, Chela basal teeth arrangement; 189. Right chela fixed finger, anterior teeth arrangement; 190. Carapace, dorsal view; 191. Right leg IV, prolateral view; 192. Right leg I, prolateral view. Scale bars: Figs 183, 184, 190–192: 0.50 mm; Figs 185-187, 189: 0.10 mm.
Remarks
This species is here removed from synonymy with G. olivaceus and re-validated on account of the distinct morphological differences between these two species, as well as the supporting molecular phylogenetic evidence.
Additional material examined
SOUTH AFRICA: Eastern Cape: 2 Adults, Port Alfred [33o 36’S, 26o 53’E], leg. J. Hewitt (AMG slide). KwaZulu-Natal: 4 Adults, 2 Immatures, Durban [29o 51’S, 31o 02’E], Mango plantation, leg. L. Bevis, 13.II.1921 (3x AMG slides); 2♀, 1♂, Same locality, leg. W.G. Rump, VIII.1940 (NMSA 648); 1♂, Same locality, Umbilo [29o 54’S, 30o 59’E], leg. L. Bevis, 27.II.1921 (NMSA 16046); 3♀, 2♂, Estcourt [29o 00’S, 29o 53’E], leg. R.F. Lawrence, VIII.1941 (NMSA 5126); 1♀, 1♂, Greytown, Mazongwaan Forest [29o 04’S, 30o 36’E], leg. R.F. Lawrance, XI.1940 (NMSA 5123); 1♀, Ixopo [30o 09’S, 30o 04’E], leg. C. F. Newson, X.1962 (NMSA 7889); 2♀, 1♂, Kranskop [28o 58’S, 30o 52’E], leg. R.F. Lawrence, I.1940 (NMSA 642); 1♀, 1 Deutonymph, Margate [30o 51’S, 30o 23’E], leg. W.G. Rump, IV.1940 (NMSA 643); 1♂, Mooi River, Rosetta [29o 18’S, 29o 59’E], leg. W.G. Rump, I.1942 (NMSA 662); 1♀, 1♂, Pietermaritzburg, [29o 36’S, 30o 23’E], leg. R. Godfrey, X.1930 (NMSA610); 8♀, 12♂, 4 immatures, Pietermaritzburg, KwaZulu-Natal National Botanical Gardens, 29o 35’S, 30o 20’E, 873 m a.s.l., Grassland, leg. M.S. Harvey, 5.IV.2001 (WAM T63156, T63157); 5♀, 4♂, Same locality, 29o 36′, 30o 21’E, 720 m a.s.l., High moisture indigenous forest, Leaf litter sifting, leg. J.A. Neethling, 23.XI.2012 (NMBA P00178); 3♀, 4♂, Pietermaritzburg, Road to Bulwer [29o 42’S, 30o 29’E], leg. R.F. Lawrence, XII.1936 (NMSA 611); 2♀, Same locality, Swartkop [29o 37’S, 30o 14’E], leg. R.F. Lawrence, VI.1957 (NMSA 5153); 2♀, 3♂, Same locality, Table Mountain [29o 37’S, 30o 36’E], leg. W.G. Rump, VII.1940 (NMSA 645); 1♀, Same locality, Town Bush [29o 36’S, 30o 23’E], leg. R.F. Lawrence, I.1940 (NMSA 5124); 1♀, Same locality, leg. R.F. Lawrence, X.1960 (NMSA 7863); 2♀, Port Edward [31o 03’S, 30o 13’E], leg. R.F. Lawrence, XII.1943 (NMSA 671); 2♀, Same locality, leg. R.F. Lawrence, VI.1944 (NMSA 675); 2♀, Port Shepstone, Oribi Gorge [30o 41’S, 30o 18’E], leg. R.F. Lawrence, X.1936 (NMSA 5125); 3♀, 5♂, Umzinto, Vernon Crookes Nature Reserve, 30o 16’S, 30o 37’E, 445 m a.s.l., High moisture indigenous forest, Leaf litter sifting, leg. J.A. Neethling, 26.XI.2012 (NMBA P00177). Western Cape: 1♀, 2♂, George, Saasveld Pass, 33o 58’S, 22o 32’E, 149 m a.s.l., Indigenous forest, Leaf litter sifting, leg. J.A. Neethling & C. Luwes, 7.XII.2012 (morphological dissection); 4♀, 3♂, Hoekwil, Woodville Big Tree, 33o 56’S, 22o 39’E, 262 m a.s.l., Indigenous forest, Leaf litter sifting, leg. J.A. Neethling & C. Luwes, 8.XII.2012 (phylogenetic analysis); 1♀, 6♂, Rheenendal, Jubilee Creek Nature Reserve, 33o 53’S, 22o 58’E, 923 m a.s.l., Afromontane forest, Leaf litter sifting, leg. J.A. Neethling & C. Luwes, 26.XII.2013 (NMBA P00173). Unknown: 2♀, No collection data (NMSA 16049).
Additional material not examined
SOUTH AFRICA: KwaZulu-Natal: 2♀, Richards Bay [28o 47’S, 32o 02’E], leg. R.F. Lawrence, XII.1943 (NMSA 670, examined by Beier 1958, not traced).
Geogarypus liomendontus sp. nov.
Figs 192-208
Holotype: ♀, SOUTH AFRICA, Limpopo, Tshwenyane, Abel Erasmus Pass, 24o 28’S, 30o 36’E, 760 m a.s.l., Indigenous mountain bush patch, Leaf litter sifting, leg. J.A. Neethling, 4.X.2012 (NMBA P00208).
Paratype: 1♂, Same data as holotype (NMBA P00209).
Etymology
The name for the species is a compound of the Greek words lioméno (λιωμένο) meaning fused, and dónti (δόντι) meaning tooth, and refers to the rows of fused basal teeth found on both chelal fingers of the female.
Diagnosis
Medium sized species. Females can reach a length (cucullus to posterior abdominal margin) of 2.11 mm, with males 1.80 mm in length. Carapace dark brown, with light brown colouration along the medial furrow and a large light brown patch on the posterior in both sexes. Pedipalp segments same colour as carapace, with the dorsal surface of the chela hand convex from the stem to the base of the chelal fingers. Abdominal tergites light brown in colour, with cream markings and indistinct dark patches in both sexes, while the abdominal sternites are either cream to light brown with very indistinct dark patches and weakly sclerotised (♀), or brown to light brown with distinct dark patches and well sclerotised (♂). All pedipalp segments with the same dark colouration as the carapace, while legs I–IV, with corresponding coxae, are cream to tan in both sexes.
This species closely resembles both G. deceptor sp. nov. and G. modjadji sp. nov. in both morphology and colouration, but can be distinguished from them by possessing rows of fused basal teeth on both chela fingers in the female. Since males of the species lack these rows of fused teeth, they remain cryptic and challenging to separate from the males of related species on chelal morphology alone, requiring examination of micro-anatomical features such as those found on the chelicerae.
Description
Carapace: Strongly sub-triangular, with narrow furrow posterior to the eyes (Fig. 206). Anterior half dark brown, light brown along medial furrow, large light brown triangular patch posteriorly, uniformly granular in both sexes. Heavily constricted anteriorly into cucullus, constriction beginning at medial furrow. Two pairs of corneate eyes situated on ocular tubercles, located about one-third away from anterior edge. Four prominent setae located on anterior edge, row of 10 setae located on posterior margin. Numerous small setae present on carapace.
Abdomen: Abdominal tergites brown to light brown, with cream markings on posterior margin of most plates, particularly those of females. Indistinct dark patches present. Tergites I and II with indistinct median dark spot each, flanked by cream patches. Tergite III without dark patches. Tergites IV–X with cream posterior margins and paired dark patches lateral of the midline. Patches may appear absent and are usually seen only on closer examination. Tergites XI and XII lack dark patches. Sternites light brown, weakly sclerotised in females; brown, well sclerotised in males. Both sexes with paired dark spots located laterally of midline. Pleural membrane wrinkled-plicate, cream.
Pedipalp: Trochanter rounded anteriorly, slightly curved, granular, with the same colouration as the carapace. Small setae located on dorsal surface, apophysis located ventrally. Pedipalp femur (Fig. 200) and patella same colour as trochanter, granular. Femur narrow at base, widening evenly until just posterior of anterior margin, where there is a sudden constriction. Pedipalp patella (Fig. 200) narrow, slightly angled at base, widening evenly into cone. Lyriform fissures visible just anterior of base, located on bulge, dorso-prolaterally on patella.
Chela: (Figs 193–196, 199, 200) Chelal hand same colour as rest of pedipalp, fingers lighter. Chela granular from stem to base of fingers. Both fingers smooth, just as long as hand (with pedicel). Dorsal surface of chelal hand uniformly convex from stem to base of chelal fingers. The prolateral surface of chela only slightly convex, retrolateral surface roughly straight. Fixed chela finger with eight trichobothria, movable chela finger with four. Both fingers long and narrow, curving to interior. Venom apparatus present on both fingers.

Figures 193–196. Digital microscope photographs of Geogarypus liomendontus sp. nov. right chela: Female (193, 194) and Male (195, 196). 193, 195. Retrolateral view; 194, 196. Dorsal view. Scale bar: 1.00 mm.
Trichobothria pattern almost identical to that of G. deceptor sp. nov., with the exception that est is located just anterior and almost opposite ib. Fixed finger with 35 teeth (♀), 38 (♂). All teeth situated in one row, several large teeth located evenly along finger length (Fig. 202). Movable finger with 27 teeth (♀), 28 (♂). Anterior edge of the female’s movable finger extends slightly beyond that of fixed chelal finger. Unique to females of this species is the presence of large fused teeth plates at the base of both chela fingers (Figs 197 & 201). Plate on fixed finger consists of fusion of first four basal teeth, teeth immediately after are tightly gouped. Plate on movable finger also consists of fusion of first four basal teeth. Males do not possess fused plates, with no increase in basal tooth size (Fig. 198).
Chelicera: Hand with five setae located as in Fig. 1. Fixed cheliceral finger with six teeth arranged as in Fig. 203. Female galea complex with nine rami (Fig. 204), simple in males (Fig. 205). Rallum composed of single blade in both sexes. Serrula exterior with 13 lamella in females, 16 in males. Lamina exterior present in both sexes.
Coxae and legs: Pedipalp coxae slightly lighter than carapace, with distinct shoulder. Legs I–IV with corresponding coxae cream to tan in both sexes. All legs diplotarsate (Figs 207 & 208).
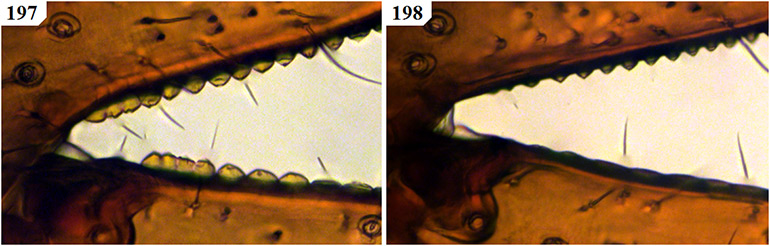
Figures 197–198. Digital microscope photographs of tooth morphology at the base of the chelal fingers of Geogarypus liomendontus sp. nov., showing distinct differences between female (197) and male (198).
Measurements: body length ♀ 2.11 ♂ 1.80; carapace ♀ 0.71 x 0.67 (1.06) ♂ 0.64 x 0.58 (1.10). Palp femur ♀ 0.71 x 0.22 (3.22) ♂ 0.65 x 0.20 (3.25), patella ♀ 0.56 x 0.21 (2.66) ♂ 0.53 x 0.20 (2.65), hand ♀ 0.61 x 0.32 ♂ 0.54 x 0.28, chela ♀ 1.22 (3.81) ♂ 1.12 (4.00), movable finger ♀ 0.59 ♂ 0.57. Leg I femur ♀ 0.31 x 0.12 ♂ 0.29 x 0.11, patella ♀ 0.19 x 0.11 ♂ 0.17 x 0.10, tibia ♀ 0.24 x 0.07 ♂ 0.23 x 0.08, metatarsus ♀ 0.17 x 0.06 ♂ 0.16 x 0.06, tarsus ♀ 0.18 x 0.04 ♂ 0.16 x 0.05. Leg IV femur ♀ 0.15 x 0.10 ♂ 0.15 x 0.10, patella ♀ 0.48 x 0.18 ♂ 0.46 x 0.18, tibia ♀ 0.40 x 0.10 ♂ 0.38 x 0.11, metatarsus ♀ 0.21 x 0.07 ♂ 0.21 x 0.07, tarsus ♀ 0.20 x 0.05 ♂ 0.20 x 0.05.
Remarks
This species is unique in that the females possess a plate consisting of large fused teeth at the base of both chelal fingers. Although G. modjadji sp. nov. females also possess fused teeth, these are present only at the base of the fixed chelal finger.
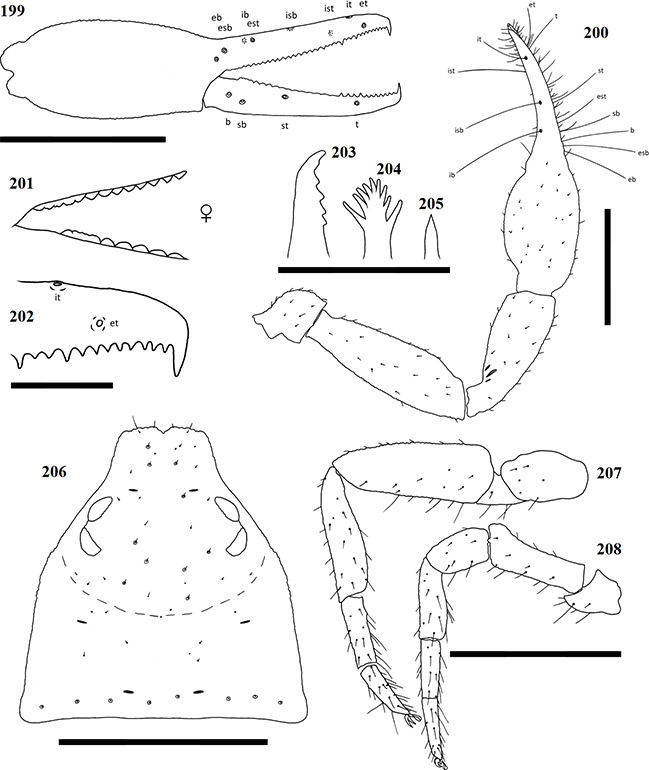
Figures 199–208. Geogarypus liomendontus sp. nov. 199–204, 206–208. Female; 205. Male. 199. Right chela, retrolateral view; 200. Right pedipalp, dorsal view; 201. Basal teeth arrangement; 202. Anterior teeth arrangement on fixed chelal finger; 203. Chelicera movable finger, dorsal view; 204, 205. Galea; 206. Carapace, dorsal view; 207. Right leg IV, prolateral view; 208. Right leg I, prolateral view. Scale bars: Figs 199, 200, 206–208: 0.50 mm; Figs 202-205: 0.10 mm.
Additional material examined
SOUTH AFRICA: Limpopo: 1♀, 1♂, Tshwenyane, Abel Erasmus Pass, 24o 28’S, 30o 36’E, 760 m a.s.l., Indigenous mountain bush patch, Leaf litter sifting, leg. J.A. Neethling, 4.X.2012 (phylogenetic analysis). Mpumalanga: 2♀, 1♂, Nelspruit, R539, 25o 24’S, 30o 44’E, 785 m a.s.l., Highveld Afromontane forest, Leaf litter sifting, leg. J.A. Neethling, 29.IX.2012 (morphological dissection).
Geogarypus modjadji sp. nov.
Figs 209–222
Holotype: ♂, SOUTH AFRICA, Limpopo, Modjadjikloof, Modjadjikloof Nature Reserve, 23o 38’S, 30o 22’E, 902 m a.s.l., Dry-shrub cycad forest, Leaf litter sifting, leg. J.A. Neethling, 5.XI.2012 (NMBA P00210).
Paratypes: 2♂, Same data as holotype (NMBA P00211).
Etymology
The species name refers to the type locality.
Diagnosis
Medium sized species. Females reach a length (cucullus to posterior abdominal margin) of 2.37 mm, while males reach 2.03 mm. Carapace dark brown to almost black, with a large cream patch on the posterior in both sexes. Pedipalp segments same colour as carapace, with the dorsal surface of the chela hand convex from the stem to the base of the chelal fingers. Abdominal tergites brown to light brown, with dark and cream patches in both sexes, while the abdominal sternites are light brown with distinct dark spots in both sexes. Pedipalp coxae lighter in colour than the carapace, with distinct shoulder, legs I–IV with corresponding coxae cream to tan in both sexes.
This species closely resembles both G. liomendontus sp. nov. and G. deceptor sp. nov. in morphology and colouration, but can be distinguished as females possess only fused teeth at the base of the fixed chela finger. Geogarypus modjadji sp. nov. is also the largest of the three species, with both sexes averaging over 2.00 mm in length. Males of G. liomendontus sp. nov. and G. deceptor sp. nov. both average below 2.00 mm in length.
Description
Carapace: Strongly sub-triangular, with narrow furrow posterior to the eyes (Fig. 220). Carapace dark brown to black, with large, triangular cream patch on posterior half, extending from posterior edge up to medial furrow. Uniformly granular in both sexes, heavily constricted anteriorly into cucullus, constriction beginning at the medial furrow. Two pairs of corneate eyes situated on ocular tubercles, located about one-third away from anterior edge. Four prominent setae located on anterior edge, row of 10 setae located on posterior margin. Numerous small setae present on carapace.
Abdomen: Abdominal tergites medium brown to light brown, dark and cream patches present on most plates. Tergites I and II with large median dark spot each, flanked by cream patches. Tergite III without dark patches, but with a large medial cream patch. Tergites IV–IX with paired dark patches just lateral of midline, flanked by cream patches, with midline also cream. Tergites IX–XII uniformly brown, lacking any discernable dark or light patches. Setae of posterior setal rows of tergites situated on small pale plates with dark edges, making these plates distinctly visible. Sternites light brown and well sclerotised in both sexes, paired dark spots located laterally of the midline. Pleural membrane wrinkled-plicate, cream.
Pedipalp: Trochanter rounded anteriorly, regular in shape, granular, with same dark colouration as carapace. Small setae located on dorsal and dorso-lateral sides, apophysis located ventrally. Pedipalp femur (Fig. 214) and patella grandular and the same colour as trochanter. Femur narrow at base, widening evenly until just posterior of anterior margin, where there is a sudden constriction. Pedipalp patella (Fig. 214) narrow, slightly angled at base, widening evenly into a cone. Lyriform fissures visible just anterior of base, located on bulge, dorso-prolaterally on patella.
Chela: (Figs 209–214) Chelal hand same colour as rest of pedipalp, fingers lighter. Chela granular from stem to base of fingers. Both fingers smooth, just as long as hand (with pedicel). Dorsal surface of chelal hand uniformly convex from stem to base of chelal fingers. Prolateral surface slightly convex, retrolateral surface roughly straight. Fixed chela finger with eight trichobothria, movable chela finger with four. Both fingers long, narrow, curving to interior. Venom apparatus present on both fingers. Trichobothrial pattern nearly identical to both G. deceptor sp. nov. and G. liomendontus sp. nov., but with the following differences: trichobothria eb and esb separated by just less than one trichobothrial width, est situated anterior of ib by one trichobothrial width. Chelal teeth acute and retrorse. Fixed finger with 38 teeth (♀), 39 (♂). All teeth situated in one row, several large teeth located equally along finger length (Fig. 216). Movable finger with 31 teeth (♀), 30 (♂). Teeth on the movable finger decrease in size into small points after about 12 teeth from anterior edge, increasing in size near base of movable finger in both sexes. Three basal teeth of fixed chelal finger of females fused (Fig. 215), those of the male grouped tightly next to each other, but not fused.

Figures 209–212. Digital microscope photographs of Geogarypus modjadji sp. nov. right chela: Female (209, 210) and Male (211, 212). 209, 211. Retrolateral view; 210, 212. Dorsal view. Scale bar: 1.00 mm.
Chelicera: Hand with five setae located as in Fig. 1. Fixed cheliceral finger with six teeth arranged as in Fig. 217. Female galea complex with nine rami (Fig. 218), simple in males (Fig. 219). Rallum composed of single blade in both sexes. Serrula exterior with 18 lamella in females, 17 in males. Lamina exterior present in both sexes.
Coxae and legs: Pedipalp coxae slightly lighter than carapace, with distinct shoulder, legs I–IV with corresponding coxae cream to tan in both sexes. All legs diplotarsate (Figs 221 & 222).
Measurements: body length ♀ 2.37 ♂ 2.03; carapace ♀ 0.77 x 0.75 (1.02) ♂ 0.67 x 0.65 (1.03). Palp femur ♀ 0.78 x 0.23 (3.39) ♂ 0.73 x 0.22 (3.31), patella ♀ 0.63 x 0.22 (2.86) ♂ 0.58 x 0.21 (2.76), hand ♀ 0.67 x 0.38 ♂ 0.62 x 0.31, chela ♀ 1.39 (3.65) ♂ 1.24 (4.00), movable finger ♀ 0.69 ♂ 0.60. Leg I femur ♀ 0.35 x 0.12 ♂ 0.32 x 0.11, patella ♀ 0.21 x 0.12 ♂ 0.19 x 0.11, tibia ♀ 0.29 x 0.09 ♂ 0.26 x 0.08, metatarsus ♀ 0.20 x 0.07 ♂ 0.18 x 0.06, tarsus ♀ 0.19 x 0.05 ♂ 0.18 x 0.05. Leg IV femur ♀ 0.15 x 0.12 ♂ 0.17 x 0.11, patella ♀ 0.53 x 0.19 ♂ 0.50 x 0.17, tibia ♀ 0.48 x 0.11 ♂ 0.42 x 0.10, metatarsus ♀ 0.26 x 0.07 ♂ 0.24 x 0.07, tarsus ♀ 0.23 x 0.06 ♂ 0.22 x 0.05.

Figures 213–222. Geogarypus modjadji sp. nov. 213–218, 220–222. Female; 219. Male. 213. Right chela, retrolateral view; 214. Right pedipalp, dorsal view; 215. Basal teeth arrangement; 216. Anterior teeth arrangement on fixed chelal finger; 217. Chelicera movable finger, dorsal view; 218, 219. Galea; 220. Carapace, dorsal view; 221. Right leg IV, prolateral view; 222. Right leg I, prolateral view. Scale bars: Figs 213, 214, 220–222: 0.50 mm; Figs 216-219: 0.10 mm.
Remarks
Currently known only from the type locality.
Additional material examined
SOUTH AFRICA: Limpopo: 3♀, 2♂, Modjadjikloof, Modjadjikloof Nature Reserve, 23o 38’S, 30o 22’E, 902 m a.s.l., Dry-shrub cycad forest, Leaf litter sifting, leg. J.A. Neethling, 5.XI.2012 (morphological dissection & phylogenetic analysis).
Geogarypus octoramosus sp. nov.
Figs 223-236
Holotype: ♀, SOUTH AFRICA, KwaZulu-Natal, Vryheid, Vryheid Mountain Nature Reserve, 27o 45’S, 30o 48’E, 1363 m a.s.l., Thicket, Leaf litter sifting, leg. J.A. Neethling & C. Luwes, 22.V.2012 (NMBA P00215).
Paratypes: 4♀, 1♂, Same data as holotype (NMBA P00216).
Etymology
The species name refers to the reduced number of eight rami on the female galea.
Diagnosis
Medium sized species. Females reach a length (cucullus to posterior abdominal margin) of 2.24 mm, while males reach 2.11 mm. Carapace medium brown, with the medial furrow and posterior edge slightly lighter in both sexes. All pedipalp segments same colour as carapace, with the dorsal surface of the chela hand convex from the stem to the base of the chelal fingers. Abdominal tergites uniformly brown, with dark patches in both sexes and paler line medially. Abdominal sternites in both sexes are brown to light brown with indistinct dark spots. Legs I–IV with corresponding coxae tan to light-yellow in both sexes.
This species closely resembles G. variaspinosus sp. nov. in general morphology and colouration, but can be easily distinguished by having eight rami on the female galea, whereas females of G. variaspinosus sp. nov. have 5–7 rami, never eight.
Description
Carapace: Strongly sub-triangular, with narrow furrow posterior to the eyes (Fig. 234). Uniformly brown with medial furrow and posterior edge slightly lighter in both sexes. Granular in both sexes, heavily constricted anteriorly into cucullus, constriction beginning at the medial furrow. Two pairs of corneate eyes situated on ocular tubercles, located about one-third away from anterior edge. Four prominent setae located on anterior edge, row of 10 setae located on posterior margin. Numerous small setae present on carapace.
Abdomen: Abdominal tergites uniformly brown, with indistinct dark patch on most plates. Tergites I and II with large median dark spot each. Tergite III lacks dark patches. Tergites IV–IX with paired dark patches just lateral of midline, tergites IX–XII lack discernible patches. Sternites of both sexes medium brown to light brown, well sclerotised. Most sternites furthermore possess paired dark spots medio-laterally. Pleural membrane wrinkled-plicate, cream.
Pedipalp: Trochanter rounded anteriorly, regular in shape, granular, with same colouration as carapace. Small setae located on dorsal and dorso-lateral sides, apophysis located ventrally. Pedipalp femur (Fig. 228) and patella same colour as trochanter, granular. Femur narrow at base, widening evenly until just posterior of anterior margin, where there is a sudden constriction. Pedipalp patella (Fig. 228) narrow and slightly angled at base, widening evenly into cone. Lyriform fissures visible just anterior of base, located on bulge, dorso-prolaterally on patella.
Chela: (Figs 223–228) Same colouration as rest of pedipalp in both sexes. Granular from stem to base of fingers. Both fingers smooth, just as long to slightly longer than hand (with pedicel). Dorsal surface of chelal hand uniformly convex from stem to base of chelal fingers. Prolateral surface convex, retrolateral surface less so. Fixed chela finger with eight trichobothria, movable chela finger with four. Both fingers long, narrow, curving to interior. Venom apparatus present on both fingers. Trichobothrial pattern nearly identical to those of G. deceptor sp. nov., G. liomendontus sp. nov. and G. modjadji sp. nov., but differs with Trichobothria b and sb positioned closer to base of movable finger. Chelal teeth acute and retrorse. Fixed finger with 34 teeth (♀), 32 (♂). Four teeth behind venom apparatus on fixed chelal finger of equal size and spacing, followed by a large tooth. Teeth situated in one row, several large teeth located equally along the finger length (Fig. 230). Movable finger with 25 teeth in both sexes. Teeth on movable finger decrease in size into small points after about 10 teeth from anterior edge, increasing in size near base of movable finger, particularly in females (Fig. 229).

Figures 223–226. Digital microscope photographs of Geogarypus octoramosus sp. nov. right chela: Female (223, 224) and Male (225, 226). 223, 225. Retrolateral view; 224, 226. Dorsal view. Scale bar: 1.00 mm.
Chelicera: Hand with five setae located as in Fig. 1. Fixed cheliceral finger with six teeth arranged as in Fig. 231. Female galea complex with eight rami (Figs 3 & 232), simple in males (Figs 4 & 233). Rallum composed of single blade in both sexes. Serrula exterior with 18 lamella in females, 17 in males. Lamina exterior present in both sexes.
Coxae and legs: Pedipalp coxae same colour as carapace, with distinct shoulder. Legs I–IV with corresponding coxae tan to light-yellow in both sexes All legs diplotarsate (Figs 235 & 236).
Measurements: body length ♀ 2.24 ♂ 2.11; carapace ♀ 0.72 x 0.76 (0.94) ♂ 0.65 x 0.64 (1.01). Palp femur ♀ 0.75 x 0.23 (3.26) ♂ 0.62 x 0.20 (3.10), patella ♀ 0.61 x 0.24 (2.54) ♂ 0.48 x 0.20 (2.40), hand ♀ 0.62 x 0.39 ♂ 0.51 x 0.32, chela ♀ 1.28 (3.28) ♂ 1.08 (3.37), movable finger ♀ 0.64 ♂ 0.55. Leg I femur ♀ 0.34 x 0.13 ♂ 0.26 x 0.11, patella ♀ 0.20 x 0.12 ♂ 0.17 x 0.10, tibia ♀ 0.26 x 0.09 ♂ 0.22 x 0.08, metatarsus ♀ 0.19 x 0.07 ♂ 0.17 x 0.06, tarsus ♀ 0.16 x 0.05 ♂ 0.16 x 0.04. Leg IV femur ♀ 0.18 x 0.11 ♂ 0.15 x 0.11, patella ♀ 0.52 x 0.19 ♂ 0.43 x 0.17, tibia ♀ 0.45 x 0.11 ♂ 0.37 x 0.11, metatarsus ♀ 0.23 x 0.07 ♂ 0.20 x 0.07, tarsus ♀ 0.23 x 0.05 ♂ 0.19 x 0.05.
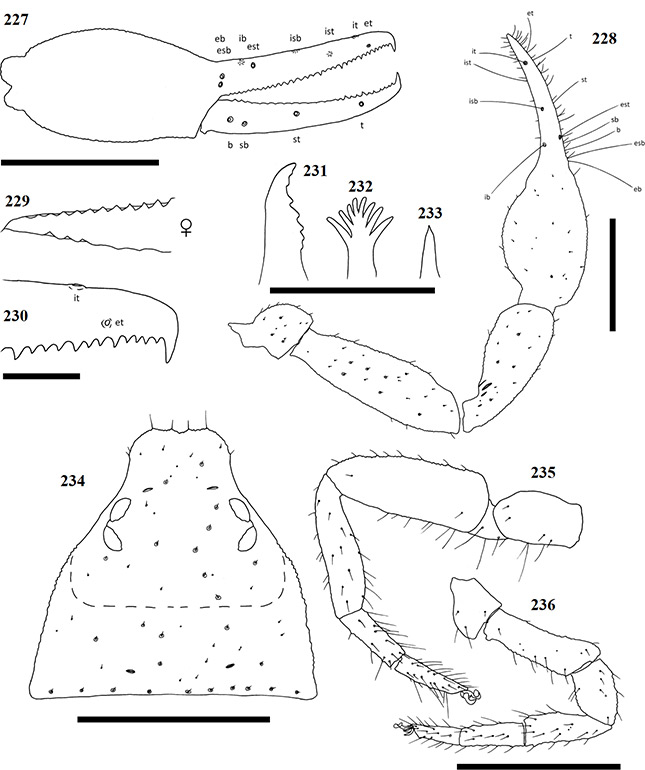
Figures 227–236. Geogarypus octoramosus sp. nov. 227–232, 234–236. Female; 233. Male. 227. Right chela, retrolateral view; 228. Right pedipalp, dorsal view; 229. Basal teeth arrangement; 230. Anterior teeth arrangement on fixed chelal finger; 231. Chelicera movable finger, dorsal view; 232, 233. Galea; 234. Carapace, dorsal view; 235. Right leg IV, prolateral view; 236. Right leg I, prolateral view. Scale bars: Figs 227, 228, 234–236: 0.50 mm; Figs 230–233: 0.10 mm.
Remarks
The reduced number of rami (eight) in females of this species was found to be constant through all populations, unlike that of G. variaspinosus sp. nov. which can vary between chelicerae of an individual.
Additional material examined
SOUTH AFRICA: Free State: 1♀, Harrismith, Platberg Nature Reserve, 28o 16’S, 29o 10’E, 1964 m a.s.l., Eucalyptus plantation, Leaf litter sifting, leg. J.A. Neethling & L. Lotz, 15.III.2012 (NMBA P00226). KwaZulu-Natal: 1♀, Emangusi, Tembe Elephant Park, 27o 02’S, 32o 25’E, 95 m a.s.l., Sand forest, leg. C.R. Haddad, 8.I.2001 (WAM T86752); 1♀, Paulpietersburg, Paulpietersburg Outskirts, 27o 20’S, 30o 29’E, 1225 m a.s.l., Grassland, Leaf litter sifting, leg. J.A. Neethling & C. Luwes, 23.V.2012 (S.E.M.); 3♀, 3♂, Vryheid, Vryheid Mountain Nature Reserve, 27o 45’S, 30o 48’E, 1363 m a.s.l., Thicket, Leaf litter sifting, leg. J.A. Neethling & C. Luwes, 22.V.2012 (morphological dissection, phylogenetic analysis and S.E.M.)
Geogarypus olivaceus (Tullgren, 1907)
Figs 237–250
Garypus olivaceus Tullgren, 1907a: 63, fig. 20 (Holotype ♀, South Africa, Free State,
Bothaville [27o 23’S, 26o 38’E], leg. H. Brauns, 9.X.1899, deposited in ZMH, examined); Tullgren, 1907b: 229.
Geogarypus (Geogarypus) olivaceus (Tullgren, 1907): Beier, 1932: 230, fig. 256; Roewer,
1937: 269; Beier, 1955: 301, fig. 20; Beier, 1958: 170.
Geogarypus olivaceus (Tullgren, 1907): Weidner, 1959: 115; Beier, 1964: 59; Beier, 1966:
461, 470; Lawrence, 1967: 89; Harvey, 1986: 760; Harvey, 1991: 258; Dippenaar-Schoeman & Harvey, 2000: 93.
Diagnosis
Medium sized species. Females can reach a length (cucullus to posterior abdominal margin) of 2.24 mm, with males reaching 2.02 mm. Carapace uniform light brown in both sexes. Pedipalp segments same colour as carapace, with the dorsal surface of the chela hand convex from the stem to the base of the chelal fingers. Abdominal tergites light brown, with dark and cream patches in both sexes, while abdominal sternites are light brown and weakly sclerotised in females, well sclerotised in males. Pedipalp coxae same colour as carapace, with distinct shoulder, legs I–IV, including coxae, yellow to tan in both sexes.
This species closely resembles smaller specimens of G. flavus stat. nov. in general morphology and colouration, which resulted in their synonymisation by Beier (1964). The two species can be separated on grounds of basal chelal teeth morphology, with G. olivaceus having large and closely spaced basal teeth (Figs 19 & 247), while G. flavus stat. nov. possess smaller and more widely spaced teeth (Figs 21 & 188).
Description
Carapace: Strongly sub-triangular, longer than broad in both sexes, with narrow furrow posterior to the eyes (Fig. 248). Carapace uniform light brown, granular in both sexes. Strongly constricted anteriorly into cucullus, constriction beginning at medial furrow. Two pairs of corneate eyes situated on ocular tubercles, located about one-third away from anterior edge. Four prominent setae located on anterior edge, row of eight setae situated along posterior edge of carapace, with small setae present and spread over carapace.
Abdomen: Abdominal tergites light brown with both dark and cream patches on most plates. Tergites I and II with large median dark spot each, flanked by cream patches. Tergite III without dark patches, but with medial cream spot. Tergites IV–X with paired dark patches just lateral of midline. Tergites IV and V with medial cream spot. Tergites XI and XII lack any patches. Sternites light brown, weakly sclerotised in females, strongly sclerotised in males. Abdomen of both sexes narrow and elongate. Pleural membrane wrinkled-plicate, cream.
Pedipalp: Trochanter rounded anteriorly, granular, with same colouration as carapace. Small setae located on dorsal and dorso-lateral sides, ventral apophysis present. Pedipalp femur (Fig. 242) and patella same colour as trochanter, granular. Femur narrow at base, widening evenly until just posterior of anterior margin, where there is a sudden constriction. Pedipalp patella (Fig. 242) narrow, slightly angled at base, widening evenly into cone. Lyriform fissures visible just anterior of base, located on bulge, dorso-prolaterally on patella.
Chela: (Figs 237–242) Same colouration as rest of pedipalp in both sexes, granular from stem to base of fingers. Both fingers smooth, just as long as hand (with pedicel). Dorsal surface of chelal hand uniformly convex from stem to base of the chelal fingers. Pro- and retrolateral surfaces convex, with prolateral surface slightly more so. Fixed chela finger with eight trichobothria, movable chela finger with four. Both fingers long, narrow, curving to interior. Venom apparatus present on both fingers. Trichobothria eb and esb situated less than one trichobothrial width from each other, retrolaterally at base of fixed finger, ib located dorso-laterally on prolateral surface, one trichobothrial width distal of esb. Trichobothria est on retrolateral surface, three trichobothrial widths distal of ib, isb situated an equal distance from est as est from ib on opposite surface. Trichobothria ist over one-third finger length from anterior finger edge, roughly in middle of prolateral surface, it on dorsal surface, one-fifth finger length from anterior finger edge. Trichobothria et just anterior of it on retrolateral surface. Trichobothria b and eb somewhat forward of finger base, b ventrally in line with esb. Trichobothria st located halfway along movable finger, t located in middle between st and anterior edge of movable finger. Chelal teeth acute and retrorse. Fixed finger with 37 teeth (♀), 35 (♂). First tooth behind the venom apparatus very small, followed by larger tooth (Fig. 246), remaining fixed chelal teeth situated in a single row, not decreasing in size basally, forming broader, less pointed teeth grouped tightly together. Movable finger with 29 teeth (♀), 28 (♂). First two teeth behind venom apparatus small and grouped close together, followed by larger teeth that decrease in size into small points. Basal teeth increase in size, grouping tightly together, almost fused in females (Fig. 247). In males, lesser degree of basal teeth size increase, teeth remaining separated.
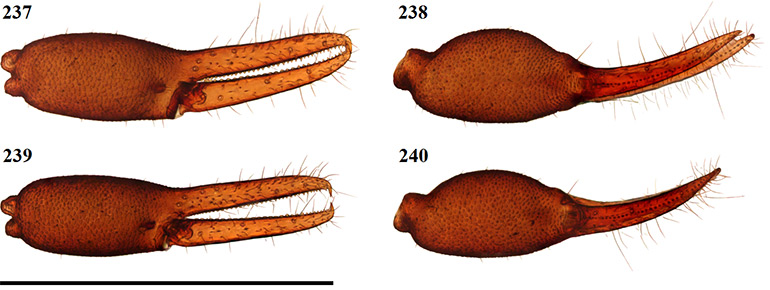
Figures 237–340. Digital microscope photographs of Geogarypus olivaceus (Tullgren, 1907). right chela: Female (237, 238) and Male (239, 240). 237, 239. Retrolateral view; 238, 240. Dorsal view. Scale bar: 1.00 mm.
Chelicera: Hand with five setae located as in Fig. 1. Fixed cheliceral finger with five teeth arranged as in Fig. 243. Female galea complex with nine rami (Fig. 244), simple in males (Fig. 245). Rallum composed of single blade in both sexes. Serrula exterior with 16 lamella in both sexes. Lamina exterior present in both sexes.
Coxae and legs: Pedipalp coxae same colour as carapace, with distinct shoulder. Legs I–IV, including coxae, yellow to tan in both sexes. All legs diplotarsate (Figs 249 & 250).
Measurements: body length ♀ 2.24 ♂ 2.02; carapace ♀ 0.61 x 0.60 (1.02) ♂ 0.58 x 0.53 (1.09). Palp femur ♀ 0.61 x 0.19 (3.21) ♂ 0.63 x 0.20 (3.15), patella ♀ 0.54 x 0.19 (2.84) ♂ 0.49 x 0.19 (2.58), hand ♀ 0.55 x 0.30 ♂ 0.52 x 0.28, chela ♀ 1.11 (3.70) ♂ 1.06 (3.78), movable finger ♀ 0.54 ♂ 0.52. Leg I femur ♀ 0.28 x 0.11 ♂ 0.29 x 0.10, patella ♀ 0.16 x 0.10 ♂ 0.16 x 0.10, tibia ♀ 0.22 x 0.06 ♂ 0.21 x 0.07, metatarsus ♀ 0.17 x 0.05 ♂ 0.16 x 0.06, tarsus ♀ 0.16 x 0.04 ♂ 0.15 x 0.04. Leg IV femur ♀ 0.13 x 0.10 ♂ 0.13 x 0.10, patella ♀ 0.44 x 0.16 ♂ 0.42 x 0.17, tibia ♀ 0.39 x 0.10 ♂ 0.37 x 0.10, metatarsus ♀ 0.22 x 0.06 ♂ 0.20 x 0.07, tarsus ♀ 0.20 x 0.05 ♂ 0.17 x 0.04.
Remarks
This was one of the first geogarypids to be described from South Africa by Tullgren (1907a). Given the morphological differences between G. olivaceus and G. flavus, together with the phylogenetic support obtained in this study, G. flavus stat. nov. is removed from synonymy with G. olivaceus and re-validated. The Western Cape distribution records for this species could potentially be mis-identifications, as no specimens were collected in the province during field work undertaken as part of this study.
Additional material examined
SOUTH AFRICA: Free State: 1♀, 1♂, Bloemfontein, Free State National Botanical Gardens, 29o 03’S, 26o 13’E, 1397 m a.s.l., Grassland, leg. C.R. Haddad, 29.I.2005 (WAM T79036, T79037); 2♀, 2♂, Same locality, Grassland, Leaf litter sifting, leg. J.A. Neethling & C. Luwes, 2.II.2012 (NMBA P00188); Bloemfontein, Maselspoort, 29o 02’S, 26o 24’E, leg. C.R. Haddad, 4.II.2005 (WAM T79035); 3♀, 1♂, Bothaville, 27o 24’S, 26o 38’E, 1262 m a.s.l., Woodland patch, Leaf litter sifting, leg. J.A. Neethling, 5.II.2012 (morphological dissection and phylogenetic analysis); 1♀, 5♂, Brandfort, Amanzi Private Game Reserve, 28o

Figures 241–250. Geogarypus olivaceus (Tullgren, 1907). 241–244, 246–250. Female; 245. Male. 241. Right chela. retrolateral view; 242. Right pedipalp, dorsal view; 243. Chelicera movable finger, dorsal view; 244, 245. Galea; 246. Right chela fixed finger, anterior teeth arrangement; 247. Female chela basal teeth arrangement; 248. Carapace, dorsal view; 249. Right leg IV, prolateral view; 250. Right leg I, prolateral view. Scale bars: Figs 241, 242, 248–250: 0.50 mm; Figs 243-246: 0.10 mm.
36’S, 26o 26’E, 1430 m a.s.l., Bushveld, Leaf litter sifting, leg. J.A. Neethling, 21.I.2014 (NMBA P00181). KwaZulu-Natal: 1♀, Newcastle, Muller’s Pass [27o 52’S, 29o 43’E], leg. R.F. Lawrence, III.1947 (NMSA 686); 1♂, Pomeroy, Gordon Memorial Mission [28o 34’S, 30o 26’E], leg. R. Godfrey, 11.IX.1930 (NMSA 16047). North West: 1♂, Brits [25o 38’S, 27o 47’E], leg. R. Watmough, 6.I.1989 (NCA 89/1005).
Geogarypus tectomaculatus sp. nov.
Figs 251–263
Holotype: ♀, SOUTH AFRICA, Mpumalanga, Graskop, Rock Formations along R532, 24o 54’S, 30o 52’E, 1500 m a.s.l., Highveld Grassland, Leaf litter sifting, leg. J.A. Neethling, 2.X.2012 (NMBA P00212).
Paratypes: 3♀, 2♂, Same data as holotype (NMBA P00213).
Etymology
The species name is a combination of the Latin words tectum meaning roof, and macula meaning spot or blemish. The name refers to the tergite markings, particularly of females, that differ from other South African geogarypid species in appearing as small distinct spots rather than large patches.
Diagnosis
Large species. Females reach an average length (cucullus to posterior abdominal margin) of 2.59 mm, while males average 2.20 mm. The carapace is dark brown in both sexes, with the posterior half being slightly lighter. Pedipalp segments are the same colour as the carapace, with the dorsal surface of the chela hand convex from the stem to the base of the chelal fingers. Abdominal tergites medium brown, with cream patches and dark spots in both sexes, while the abdominal sternites are medium brown to light brown with dark spots in both sexes. Legs I–IV, including coxae, tan to pale yellow in both sexes.
Closely resembling G. flavus stat. nov., G. tectomaculatus sp. nov. can be distinguished by having fewer chelal teeth as well as possessing relatively large basal teeth that are grouped tightly together, while G. flavus stat. nov. has smaller basal teeth that are separate from one another.
Description
Carapace: Strongly sub-triangular, slightly longer than broad in both sexes, with narrow furrow posterior to eyes (Fig. 261). Posterior half somewhat lighter than anterior half. Heavily constricted anteriorly into cucullus, constriction beginning at medial furrow. Two pairs of corneate eyes situated on ocular tubercles, located about one-third from anterior edge. Four prominent setae located on anterior edge, row of 10 setae located on posterior margin. Numerous small setae present on carapace.
Abdomen: Abdominal tergites brown with cream patches and dark spots on most plates. Tergites I and II with median dark spot each, flanked by cream patches. Tergite III without dark patches, but with central cream coloured band extending almost entire length of tergum. Tergites IV–X with medial cream patch, with paired dark spots just lateral of midline, flanked laterally by small cream patches. Dark spots distinctly smaller in females, sometimes less distinct and larger in males. Tergites XI and XII lack patches. Setae of posterior setal rows of tergites, predominantly in females, situated on pale plates with dark edges, making plates distinctly visible. Sternites well sclerotised in both sexes, with paired dark points located laterally of cream medial line. Pleural membrane wrinkled-plicate, cream.
Pedipalp: Trochanter rounded anteriorly, granular, with same colouration as carapace. Small setae located on dorsal and dorso-lateral sides, ventral apophysis present. Pedipalp femur (Fig. 256) and patella granular, same colour as trochanter. Femur narrow at base, widening evenly until just posterior of anterior margin, where it is suddenly constricted. Pedipalp patella (Fig. 256) narrow, slightly angled at base, widening evenly into cone. Lyriform fissures visible just anterior to base, located on bulge, dorso-prolaterally on patella.
Chela: (Figs 251–256) Same colour as rest of pedipalp in both sexes, granular from stem up to base of fingers. Both fingers smooth, just as long as hand (with pedicel). Dorsal surface of chelal hand uniformly convex in shape from stem to base of chelal fingers. Both pro- and retrolateral surfaces slightly convex in shape, prolateral surface more so than retrolateral. Fixed chela finger with eight trichobothria, movable chela finger with four trichobothria. Both fingers long, narrow, curving to interior. Venom apparatus present on both fingers. Trichobothria eb and esb situated just under one trichobothrial width from each other, located retrolaterally at base of fixed finger, ib located dorso-laterally on prolateral surface, four trichobothrial widths distal of esb. Trichobothria est located one trichobothrial width distal, opposite from ib in females, three trichobothrial widths distal in males, resulting in est positioned just proximal of isb. Trichobothria isb located almost dorsally, roughly halfway along finger in females, one-third finger length from base of fixed finger in males, while ist located one-third finger length from anterior finger edge, roughly in middle of the prolateral surface in both sexes. Trichobothria it located on dorsal surface, roughly one-quarter finger length from anterior finger edge, et situated anteriorly on retrolateral surface. Trichobothria b and eb on movable finger situated somewhat forward of finger base, b ventrally in line with esb. Trichobothria st positioned just before halfway along movable finger, t located midway between st and anterior edge of movable finger. Chelal teeth acute, retrorse, curving back slightly. Fixed finger with 34 teeth (♀), 36 (♂). First three teeth behind venom apparatus small, grouping together to form distinct group (Fig. 260). This group is followed by remaining fixed chelal teeth, situated in a single row, only decreasing slightly in size to base, with distinctly larger teeth located throughout length of fixed finger. Movable finger with 28 teeth (♀), 30 (♂). In both sexes basal teeth on movable finger increase in size into distinct teeth, following small tooth points.
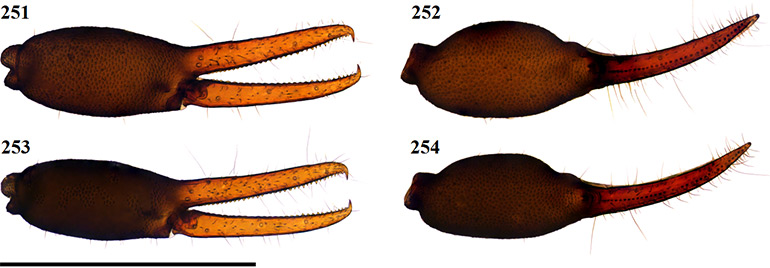
Figures 251–254. Digital microscope photographs of Geogarypus tectomaculatus sp. nov. right chela: Female (251, 252) and Male (253, 254). 251, 253. Retrolateral view; 252, 254. Dorsal view. Scale bar: 1.00 mm.
Chelicera: Hand with five setae located as in Fig. 1. Fixed cheliceral finger with six teeth arranged as in Fig. 257. Female galea complex with nine rami (Fig. 258), simple with single spinule in males (Fig. 259). Rallum composed of single blade in both sexes. Serrula exterior with 15 lamella in both sexes. Lamina exterior present in both sexes.
Coxae and legs: Pedipalpal coxae same colour as carapace, with distinct shoulder. Legs I–IV, including coxae, tan to pale yellow in both sexes. All legs diplotarsate (Figs 262 & 263).
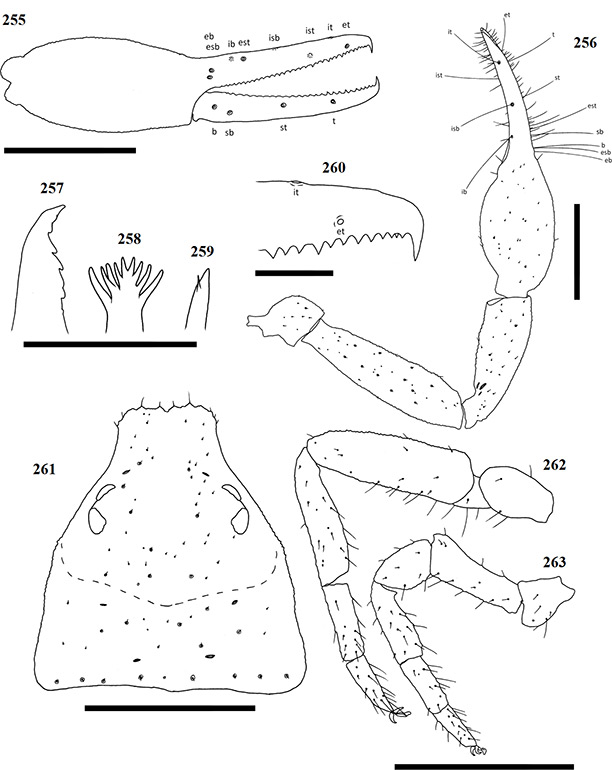
Figures 255–263. Geogarypus tectomaculatus sp. nov. 255–258, 260–263. Female; 259. Male; 255. Right chela, retrolateral view; 256. Right pedipalp, dorsal view; 257. Chelicera movable finger, dorsal view; 258, 259. Galea; 260. Right chela fixed finger, anterior teeth arrangement; 261. Carapace, dorsal view; 262. Right leg IV, prolateral view; 263. Right leg I, prolateral view. Scale bars: Figs 255, 256, 261–263: 0.50 mm; Figs 257–260: 0.10 mm.
Measurements: body length ♀ 2.59 ♂ 2.20; carapace ♀ 0.81 x 0.80 (1.01) ♂ 0.77 x 0.73 (1.05). Palp femur ♀ 0.88 x 0.24 (3.66) ♂ 0.81 x 0.23 (3.52), patella ♀ 0.70 x 0.24 (2.91) ♂ 0.67 x 0.23 (2.91), hand ♀ 0.72 x 0.39 ♂ 0.68 x 0.34, chela ♀ 1.45 (3.71) ♂ 1.41 (4.14), movable finger ♀ 0.71 ♂ 0.72. Leg I femur ♀ 0.37 x 0.13 ♂ 0.34 x 0.13, patella ♀ 0.21 x 0.12 ♂ 0.20 x 0.12, tibia ♀ 0.29 x 0.09 ♂ 0.28 x 0.09, metatarsus ♀ 0.20 x 0.07 ♂ 0.19 x 0.07, tarsus ♀ 0.18 x 0.06 ♂ 0.17 x 0.05. Leg IV femur ♀ 0.17 x 0.12 ♂ 0.17 x 0.11, patella ♀ 0.56 x 0.18 ♂ 0.53 x 0.19, tibia ♀ 0.46 x 0.12 ♂ 0.46 x 0.11, metatarsus ♀ 0.26 x 0.08 ♂ 0.25 x 0.07, tarsus ♀ 0.25 x 0.06 ♂ 0.24 x 0.06.
Remarks
This is the second largest geogarypid species in South Africa, being just slightly smaller than Afrogarypus robustus comb. nov. Males of the species also possess the largest chela ratios, with chela that are more than four times as long as wide.
Additional material examined
SOUTH AFRICA: Mpumalanga: 1♀, Graskop, God’s Window, 24o 52’S, 30o 53’E, 1705 m a.s.l., Highveld bush with indigenous forest patches, Leaf litter sifting, leg. J.A. Neethling, 28.IX.2012 (morphological dissection); 3♀, 1♂, Same locality, Wonderview, 24o 52’E, 30o 54’E, 1723 m a.s.l., Highveld bush, Leaf litter sifting, leg. J.A. Neethling, 30.IX.2012 (morphological dissection & phylogenetic analysis); 1♂, Sabie, Bergvliet State Forest, 25o 10’S, 30o 48’E, Pitfall trapping, leg. A. van den Berg, 27.XI.1986 (NCA 87/444).
Geogarypus variaspinosus sp. nov.
Figs 264–278
Holotype: ♀, SOUTH AFRICA, Northern Cape, Olifantshoek, 27o 56’S, 22o 44’E, 1294 m a.s.l., Rocky veld, Leaf litter sifting, leg. J.A. Neethling, 16.VI.2013 (NMBA P00223).
Paratypes: 2♂, Same data as holotype (NMBA P00225).
Etymology
The species name refers to the variable number of rami that occur on the galea of a female.
Diagnosis
Medium sized species. Females reach a length (cucullus to posterior abdominal margin) of 2.05 mm, while males reaching 1.99 mm. Carapace overall medium brown, with the medial furrow and posterior edge slightly lighter in both sexes. All pedipalp segments same colour as carapace, with the dorsal surface of the chela hand convex from the stem to the base of the chelal fingers. Abdominal tergites uniformly brown to light brown, with dark patches in both sexes, while the abdominal sternites are light brown with indistinct dark spots. Legs I–IV, including coxae, tan in both sexes.
This species closely resembles Geogarypus octoramosus sp. nov. in general morphology and colouration, but can be distinguished by the variable number of rami found on the galea of females, which are always fewer than eight (5–7). Males are difficult to distinguish from related congeners, but resemble smaller, lighter females.
Description
Carapace: Strongly sub-triangular, longer than broad in both sexes, with narrow furrow posterior to eyes. Carapace uniformly brown, granular, furrow and posterior edge slightly paler. Heavily constricted interiorly into cucullus, constriction beginning at medial furrow. Two pairs of corneate eyes situated on ocular tubercles, located about one-third away from anterior edge (Fig. 276). Four prominent setae located on anterior edge, row of 10 setae located on posterior margin.
Abdomen: Abdominal tergites overall light brown, dark patches and indistinct cream markings present. Tergites I and II with large medial dark spot each. Tergite III lacks dark patches. Tergites IV–X with paired dark patches just lateral of the midline. May appear very indistinct in some specimens. Tergites XI and XII lack dark patches. Sternites light brown, well sclerotised in both sexes. Paired dark spots, located laterally of midline, present. Pleural membrane wrinkled-plicate, cream.
Pedipalp: Trochanter rounded anteriorly, slightly curved, granular, same colour as carapace. Small setae located on dorsal and dorso-lateral sides, ventral apophysis present. Pedipalp femur (Fig. 269) and patella same colour as trochanter, granular. Femur narrow at base, widening evenly until just posterior of anterior margin, where it is constricted. Pedipalp patella (Fig. 269) narrow and slightly angled at base, widening evenly into cone. Lyriform fissures visible just anterior of base, located on a bulge, dorso-prolaterally on patella.
Chela: (Figs 264–269) Same colouration as rest of pedipalp in both sexes, granular from stem up to base of fingers. Both fingers smooth, as long to slightly shorter than hand (with pedicel). Dorsal surface of chelal hand uniformly convex in shape from the stem to the base of the chelal fingers. Both pro- and retrolateral surfaces slightly convex in shape, prolateral surface slightly more so. Fixed chela finger with eight trichobothria, movable chela finger with four trichobothria. Both fingers long and narrow, curving to the interior. Venom apparatus present on both fingers.

Figures 264–267. Digital microscope photographs of Geogarypus variasponosum sp. nov. right chela: Female (264, 265) and Male (266, 267). 264, 266. Retrolateral view; 265, 267. Dorsal view. Scale bar: 1.00 mm.
Trichobothria eb and esb situated one trichobothrial width apart, located retrolaterally at base of fixed finger, ib located dorso-prolaterally, four trichobothrial widths distal of esb. Trichobothria est located on retrolateral surface, just anterior of ib, isb situated an equal distance from est as est is from esb, on opposite surface. Trichobothria ist located one-third finger length from anterior finger edge, somewhat ventral on prolateral surface, it located on dorsal surface, one-fifth finger length from anterior finger edge. Trichobothria et situated just anterior of it, on retrolateral surface. Trichobothria b and eb are situated somewhat forward of finger base, b ventrally in line with esb. Trichobothria st variable, positioning from one-third finger length from base of finger to just under halfway along length of movable finger. Trichobothria t is located one-quarter finger length from anterior edge of movable finger. Chelal teeth acute, retrorse, some curving back slightly. Fixed finger with 38 teeth (♀), 40 (♂). First two teeth behind venom apparatus small, followed by larger tooth (Fig. 271). Rest of fixed finger teeth situated in single row, not decreasing in size to base of finger. In females basal seven teeth are fused, followed by teeth that group tightly next to each other. Movable finger with 31 teeth (♀), 32 (♂). First two teeth behind the venom apparatus of movable finger small, grouped closely together, followed by larger teeth that reduce in size posteriorly into tooth plates. In females basal teeth increased in size and group tightly together, seeming almost fused (Fig. 270). In males there is a lesser degree of basal teeth regeneration, teeth remaining widely apart.
Chelicera: Hand with five setae located as in Fig. 1. Fixed cheliceral finger with six teeth arranged as in Fig. 272. Female galea complex with variable number of rami, ranging from five to seven (Figs 273 & 274). Male galea simple, without spinules (Fig. 275). Rallum composed of a single blade in both sexes. Serrula exterior with 16 lamella in both sexes. Lamina exterior present in both sexes.
Coxae and legs: Pedipalpal coxae same colour as carapace, with distinct shoulder. Legs I–IV, including coxae, tan in colour in both sexes. All legs diplotarsate (Figs 277 & 278).
Measurements: body length ♀ 2.05 ♂ 1.99; carapace ♀ 0.78 x 0.74 (1.05) ♂ 0.68 x 0.66 (1.03). Palp femur ♀ 0.80 x 0.24 (3.33) ♂ 0.71 x 0.20 (3.55), patella ♀ 0.67 x 0.24 (2.79) ♂ 0.59 x 0.20 (2.95), hand ♀ 0.63 x 0.36 ♂ 0.55 x 0.29, chela ♀ 1.30 (3.61) ♂ 1.08 (3.72), movable finger ♀ 0.64 ♂ 0.52. Leg I femur ♀ 0.34 x 0.13 ♂ 0.32 x 0.12, patella ♀ 0.19 x 0.11 ♂ 0.16 x 0.10, tibia ♀ 0.28 x 0.08 ♂ 0.24 x 0.08, metatarsus ♀ 0.18 x 0.06 ♂ 0.17 x 0.05, tarsus ♀ 0.16 x 0.05 ♂ 0.16 x 0.04. Leg IV femur ♀ 0.17 x 0.11 ♂ 0.17 x 0.11, patella ♀ 0.55 x 0.20 ♂ 0.52 x 0.19, tibia ♀ 0.47 x 0.11 ♂ 0.45 x 0.11, metatarsus ♀ 0.24 x 0.07 ♂ 0.22 x 0.07, tarsus ♀ 0.24 x 0.06 ♂ 0.22 x 0.06.
Remarks
Geogarypus variaspinosus sp. nov. is unique in that all females studied possessed chelicerae with galeae that differed in the number of rami present. Of particular interest is that in many specimens the number of rami varied even between the left and right sides, but always numbered from five to seven.
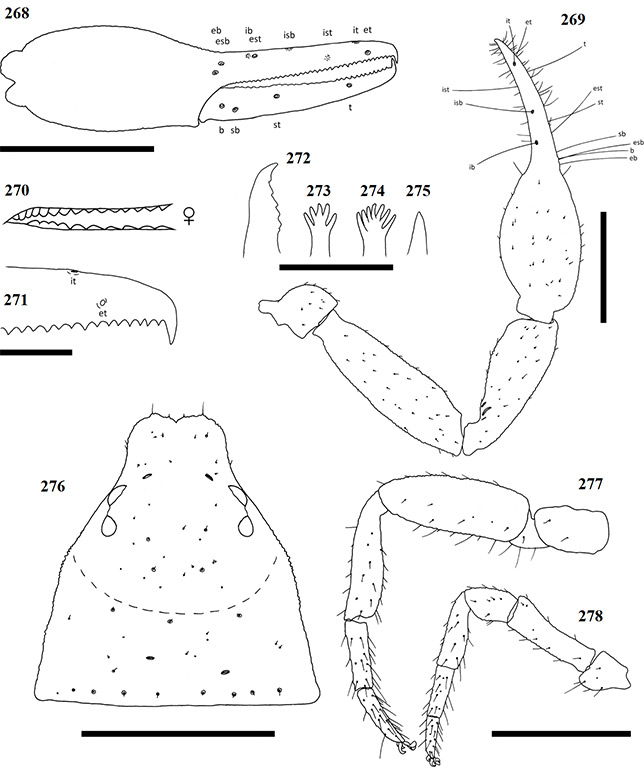
Figures 268–278. Geogarypus variaspinosus sp. nov. 268–274, 276–278. Female; 275. Male. 268. Right chela, retrolateral view; 269. Right pedipalp, dorsal view; 270. Basal teeth arrangement; 271. Anterior teeth arrangement on fixed chelal finger; 272. Chelicera movable finger, dorsal view; 273, 274, 275. Galea; 276. Carapace, dorsal view; 277. Right leg IV, prolateral view; 278. Right leg I, prolateral view. Scale bars: Figs 268, 269, 276–278: 0.50 mm; Figs 271–275: 0.10 mm.
Additional material examined
SOUTH AFRICA: Northern Cape: 1♀, 1♂, Douglas, Sunset View River Guesthouse, 29o 04’S, 23o 44’E, 985 m a.s.l., Bushveld, Leaf litter sifting, leg. J.A. Neethling, 19.VI.2013 (NMBA P00224); 4♀, 4♂, Olifantshoek, 27o 56’S, 22o 44’E, 1294 m a.s.l., Rocky veld, Leaf litter sifting, leg. J.A. Neethling, 16.VI.2013 (morphological dissection and phylogenetic analysis).
Biogeography
Worldwide, the Geogarypidae are now represented by 76 recognized species recorded from 69 countries, distributed mostly along the equatorial belt (Petrov 1997; Harvey 2000, 2013). They can be found in habitats ranging in altitude from 0–3200 m above sea level (Beron 2002). South Africa currently has the highest number (18) of described species, followed by Kenya (8) and India (7). The family also contains one of the most widespread pseudoscorpion species, Geogarypus longidigitatus (Rainbow, 1897), which can be found across 150o of longitude, throughout the Pacific region from Thailand to Easter Islands (Harvey 2000). Predominantly tropical and temperate in nature, most Geogarypidae dwell in leaf litter in forested areas. Their adaptability has allowed some species to inhabit more peculiar habitats, including intertidal zones (Haddad & Dippenaar-Schoeman 2009) and birds’ nests (Turienzo, Di Iorio & Mahnert 2010).
Within the Geogarypidae, the genus Geogarypus is the most widespread, with species being recorded from southern Europe, Africa, Asia, Australia, and as far south as Argentina in South America (Harvey 1981, 1986, 1987b; Mahnert et al. 2011). The remaining genera, on the other hand, are more restricted in their distribution, with Afrogarypus recorded only from Africa and surrounding islands such as the Seychelles (Spaull 1979), and Indogarypus being restricted to India and Sri Lanka (Harvey 1986).
The climate in South Africa is characterised by increasing aridity in an east to west plane, with the highest precipitation occurring in the coastal regions along mountain ranges. These areas regularly receive annual rainfall above 1000 mm. Some of the most western parts of the subcontinent receive less than 250 mm annually, while the interior generally varies in rainfall from 300–800 mm annually (Mansell & Erasmus 2002). Two of the world’s floristic regions can be found within South Africa, namely the Paleotropical Region (comprising mainly open woodland, savanna and grassland vegetation) and the Cape Floristic Region (comprising fynbos vegetation), with the latter being endemic to the southern part of South Africa (Cox 2001). Mucina & Rutherford (2006) recognized eight distinct biomes within South Africa, namely Albany Thicket, Desert, Indigenous Forest, Fynbos, Grassland, Nama-Karoo, Savanna and Succulent Karoo (Fig. 279).
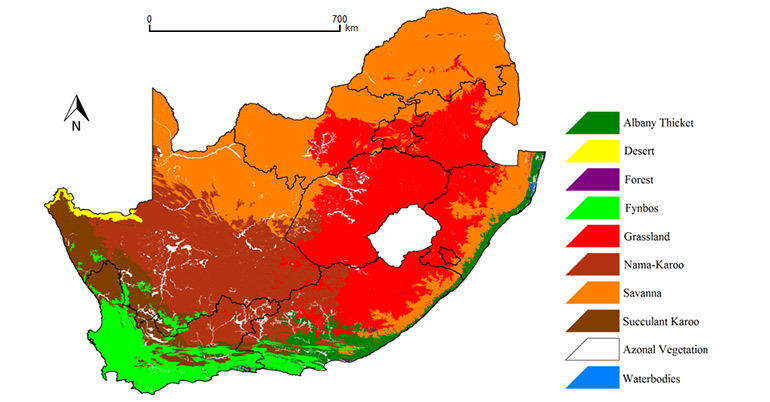
Figure 279. Vegetation map of South Africa, depicting the biomes as delineated by Mucina & Rutherford (2006). Vegetation map shapefile acquired from http://bgis.sanbi.org/vegmap/biomes.asp (accessed 12.X.2014).
In terms of biodiversity, particular emphasis is placed on the afore-mentioned Cape Floristic Region (CFR) hotspot and the Maputaland-Pondoland-Albany (MPA) hotspot (Fig. 280). The latter is the second richest floristic region after the CFR and incorporates six of the eight biomes in South Africa. It covers an area of approximately 275 000 km2 below the Great Escarpment along the eastern coast, extending to the coastal areas of central Mozambique (Perera, Ratnayake-Perera & Procheş 2011).
Species biogeography and ecology
When studying the distribution of the South African Geogarypidae (Fig. 280), it is immediately apparent that there is a noticeable north-south division in the distribution of records of Afrogarypus and Geogarypus, with the first predominantly occurring in the southern part of the country and the latter in the northern part. Distribution of the genera is furthermore concentrated along the higher rainfall areas found on the coastal side of the mountain ranges, especially within the fynbos, Albany thicket and indigenous forest biomes.
The large gaps located in the interior of the country are possibly due to poor sampling, particularly in the Northern Cape and North West provinces. When comparing Fig. 280 to Griswold (1991) and Haddad (2014), many of the groupings formed by the Geogarypidae fall perfectly within three known areas of endemism, which are the CFR and the Knysna indigenous forests, both in southern South Africa, and the MPA centre of endemism, which extends from south-eastern South Africa to central Mozambique. When comparing the distribution of individual species (Figs 281–298), one can see that many fall entirely within these centres of endemism. Other species such as A. impressus and A. minutus comb. nov. have distribution ranges that cover large areas of southern and south-eastern South Africa, although some of the records are currently unconfirmed and may prove to be mis-identifications.
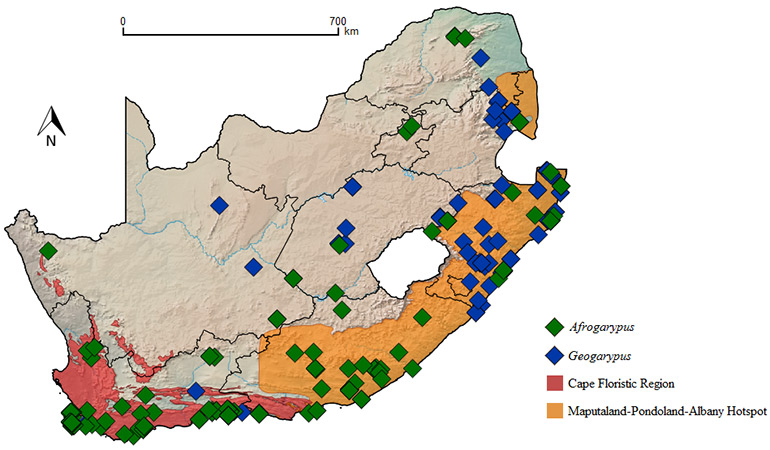
Figure 280. Topographical map of South Africa displaying the distribution of the two Geogarypidae genera in South Africa, illustrating a noticeable north-south division between the distribution of Afrogarypus Beier, 1931 and Geogarypus Chamberlin, 1930 records. Also highlighted are the distributions of the Cape Floristic Region (CFR) and the Maputaland–Pondoland–Albany (MPA) hotspot. Shapefile acquired from http://www.conservation.org/where/priority_areas/hotspots/Pages/hotspots_main.aspx (accessed 12.X.2014).
Afrogarypus carmenae sp. nov.
Distributed entirely within the CFR (Fig. 281), A. carmenae sp. nov. can be found in the leaf litter of mesic mountain fynbos within the central parts of the Western Cape, and in leaf litter of coastal fynbos around the southern edges of the province. Most exemplars were collected from the Cape of Good Hope Nature Reserve, though never in high numbers. In contrast, those collected from the inland Cederberg Mountain areas near Clanwilliam were found in greater numbers in deep leaf litter collected amongst rocks under dense shrubs. Specimen records indicate that the species can be collected throughout the year.
Habitat & Biology: Leaf litter of mesic mountain and coastal fynbos, particularly from deep leaf litter that is only slightly moist. Elevation: 7–334 m.
Afrogarypus castigatus sp. nov.
This species has so far been collected only from coastal forests and wetlands in the southern part of the iSimangaliso Wetland Park, located in the central part of the MPA centre of endemism in south-eastern Maputaland (Fig. 282). The species is furthermore significant in that it appears to be the only obligate arboreal geogarypid in South Africa, collected only from the canopies of indigenous trees within its distribution range. Geogarypus deceptor sp. nov. occurs within the same area as A. castigatus sp. nov., but was found only in leaf litter, while A. castigatus sp. nov. was never collected from leaf litter. It is therefore considered to be a short-range endemic to the area.
Habitat & Biology: To date, found under the bark and in crevices of the trees Breonadia salicina (Matumi), Trichilia emetica (Natal Mahogany) and Syzygium cordatum (Umdoni Waterberry). The only specimens collected thus far were sampled during May. Elevation: 14–24 m.
Afrogarypus excelsus (Beier, 1964) stat. nov.
This species is limited to the south-central region of the Eastern Cape (Fig. 283), with its distribution falling completely within the MPA centre of endemism. The region is characterised by its myriad of isolated mountains and gorges, each with accompanying indigenous forest patches. It is within these forest patches, as well as surrounding vegetation, where A. excelsus stat. nov. is most often found. No specimens have been collected from the grasslands between these forest fragments, and as such it is unclear whether populations are confined to these forests or not.
Habitat & Biology: Leaf litter of indigenous high moisture Afromontane forests and dense vegetation stands, particularly around rock formations. All exemplars were collected between November to March. Elevation: 500–1200 m.
Afrogarypus impressus (Tullgren, 1907)
Preferring deep, moist leaf litter, A. impressus occurs throughout the Afromontane forests along the coastal provinces, but has also been recorded from more arid regions in the Northern Cape and Mpumalanga. Its distribution is not confined to South Africa, and the species has been reported from as far north as the coral atoll of the outer Seychelles island of Aldabra in the Indian Ocean (Spaull 1979). The species has a mostly coastal distribution (Fig. 284) with few inland records, with the exception of the unconfirmed records from the Van Reenen area in the Drakensberg Mountains, which may be mis-identifications of G. octoramosus sp. nov., which has been collected from the nearby Platberg Nature Reserve. A large proportion of localities fall within both the Knysna and Tsitsikamma indigenous forests, contained within the CFR, with several also located within the MPA centre of endemism in the Eastern Cape and KwaZulu-Natal. Since its initial collection from leaf litter in the Amanzimtoti region on the 18 January 1905 (Tullgren 1907b), the species has become one of the most extensively recorded geogarypid species in South Africa. Most localities fall within the 0–300 m elevation range.
Habitat & Biology: Leaf litter of indigenous high moisture Afromontane forests along the South African coast, particularly around rock formations. Found under rocks in more arid regions in the Northern Cape and savannas in Mpumalanga. Elevation: 50–1600 m. Of special interest is that Ellingsen (1912) published notes he received from the Rev. B. Godfrey on parts of the life history of the species. This makes A. impressus the only South African geogarypid with published biological data. According to the notes the species is ground-living and found under stones in forests. Its nests are attached to stones, hemispherical in shape and lined with both silk and ground particles. Moulting has been recorded in the months of April and September, with females both gravid and with brood-sacs recorded in December (Ellingsen 1912).
Afrogarypus megamolaris sp. nov.
Known from only a handful of localities, including one that is within the Transvaal-Drakensberg endemism area, as proposed by Griswold (1991). The species’ distribution spans three provinces (Fig. 285), with all localities located within the Savanna Biome. Specimens tend to be difficult to find as they seem to prefer deep, moist, shaded and compact leaf litter found alongside large rock formations within the drier northern savanna.
Habitat & Biology: Deep, moist and compact leaf litter, especially next to large shaded rock formations, such as those of the Soutpansberg Mountains. Specimens have all been collected during the warmer months of September to March. Elevation: 300–1400 m.
Afrogarypus minutus (Tullgren, 1907) comb. nov.
Members of this species have been recorded primarily along the coastal areas of the Western Cape, Eastern Cape and KwaZulu-Natal, with some of the distribution records being within both the CFR and MPA centres of endemism (Fig. 286). Specimens have been sampled from leaf litter mainly in Coastal Fynbos and Albany Thicket, but also in indigenous forest.
Habitat & Biology: Moist and dry leaf litter from Fynbos and Albany Thicket shrubs, primarily in coastal areas. Specimens have been recorded throughout the year, although most were sampled during December. Elevation: 50–2100 m.
Afrogarypus purcelli (Ellingsen, 1912) comb. nov.
Mostly occurring within the southern coastal region of the Western Cape, sympatrically with A. carmenae sp. nov. (Fig. 287), this species has nonetheless also been recorded from the Tembe Elephant Park in KwaZulu-Natal, with unconfirmed reports from Makhado (Louis Trichardt) in Limpopo. It is one of the most widely distributed South African species. The reason for the species’ absence in the large area between the CFR and its
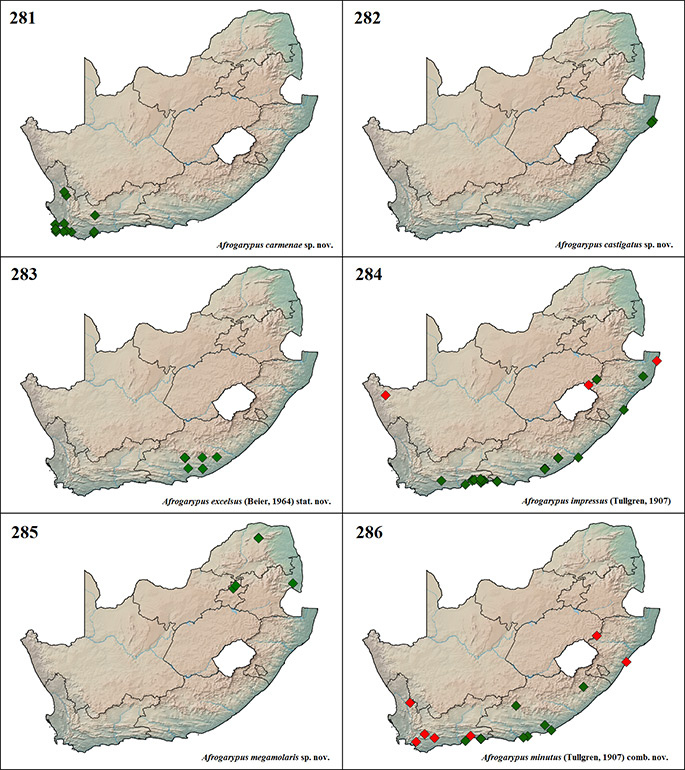
Figures 281–286. Geographical distribution of: 281. Afrogarypus carmenae sp. nov.; 282. A. castigatus sp. nov.; 283. A. excelsus (Beier, 1964) stat. nov.; 284. A. impressus (Tullgren, 1907); 285. A. megamolaris sp. nov.; 286. A. minutus (Tullgren, 1907) comb. nov., where green indicates confirmed localities and red unconfirmed published data.
northern distribution is unclear at the moment. It is possible, however, that the specimens collected from Louis Trichardt in Limpopo that Beier (1964) attributed to this species could be mis-identifications of a yet unknown species, as the location falls within the Transvaal-Drakensberg endemism area.
Habitat & Biology: Western Cape exemplars were collected during December from leaf litter of the same mesic mountain and coastal fynbos as A. carmenae sp. nov. Specimens from the northern areas were collected from leaf litter in savanna. Elevation: 40–950 m.
Afrogarypus robustus (Beier, 1947) comb. nov.
A short-range endemic species from the CFR. All known specimens were collected from either indigenous or mixed-indigenous forest patches on and around Table Mountain (Fig. 288). The species is therefore entirely restricted to the Table Mountain area of endemism, as proposed by Griswold (1991). This is the third pseudoscorpion species restricted to Table Mountain, in addition to Chthoniella cavernicola (Lawrence, 1935) and Gymnobisium
inukshuk Harvey & Giribet, 2016 (Picker & Samways 1996; Harvey et al. 2016).
Habitat & Biology: Moist or dry leaf litter next to logs and rock formations in indigenous forests located primarily within gorges on Table Mountain, such as Skeleton Gorge at the Kirstenbosch National Botanical Gardens. All specimens have been sampled from the months of August to January. Elevation: 50–900 m.
Afrogarypus subimpressus (Beier, 1955)
Originally only known from its type locality (Beier 1955), the distribution of the species has since increased to include regions of the eastern Western Cape, adjoining Northern Cape and southern Free State (Fig. 289). Primarily collected from the Cape Point Nature Reserve in the Fynbos Biome of the CFR, the species can also be found in leaf litter under shrubs in grassland areas located further inland.
Habitat & Biology: Primarily in moist leaf litter under fynbos vegetation, particularly adjacent to coastal rock formations. Specimens have been recorded from April to December, with most specimens sampled from October to December. Elevation: 10–1400 m.
Afrogarypus triangularis (Ellingsen, 1912) comb. nov.
Although occurring primarily in the Eastern Cape, this species has also been recorded from the Free State (Fig. 290). Semi-arboreal in nature, A. triangularis was collected in canopy fogging samples taken from Podocarpus falcatus (Yellowwood) and other indigenous trees found in forests throughout the Eastern Cape, where they have also been reported from bird nests (Ellingsen 1912). Individuals may also be collected from moist leaf litter in these areas. In grasslands the species can be found in grass tussocks and shaded leaf litter. Interestingly, most locations fall either within the 600–700 m or 1200–1400 m elevation ranges.
Habitat & Biology: Semi-arboreal species preferring canopies of indigenous trees in forest stands in the Eastern Cape, as well as associated leaf litter; and found in shaded leaf litter and grass tussocks in grassland habitats. Elevation: 60–1485 m.
Geogarypus deceptor sp. nov.
Occurring mainly within the north and north-eastern parts of the MPA centre of endemism, this species can be found amongst leaf litter in habitats ranging from coastal forests to wetlands, closed woodlands and savanna. The species occurs mainly in the northern parts of KwaZulu-Natal, extending to the centre of Mpumalanga (Fig. 291).
Habitat & Biology: Specimens have been collected primarily in dry leaf litter in coastal forests and woodland areas, but have also been sampled from very moist leaf litter in the indigenous forest patch located at the Highveld Botanical Gardens in Nelspruit. Recorded throughout the year. Elevation: 30–620 m.
Geogarypus flavus (Beier, 1947) stat. nov.
The distribution of this species is split between two main areas (Fig. 292), with a single locality record between them. The southernmost population falls within the Knysna indigenous forest area of endemism, while the northern population is contained within the MPA centre of endemism in KwaZulu-Natal. A common species of the coastal and Afromontane forest belts; most specimens were collected from high moisture leaf litter in indigenous forest patches.
Habitat & Biology: High moisture leaf litter from indigenous forest patches, particularly deep, loose litter next to large rocks. Specimens have been recorded throughout the year, with most sampled during the summer months of December to February. Elevation: 10–1100 m.
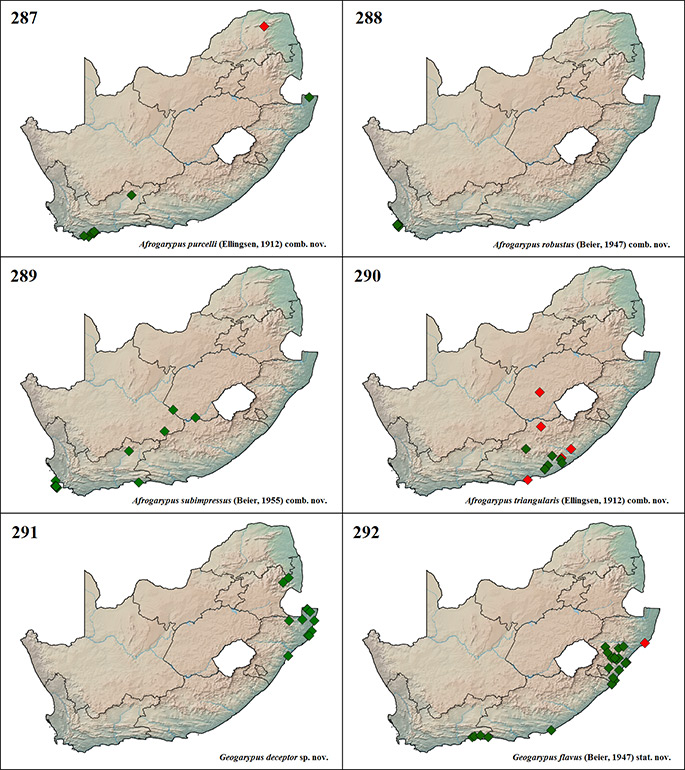
Figures 287–292. Geographical distribution of: 287. Afrogarypus purcelli (Ellingsen 1912) comb. nov.; 288. A. robustus (Beier, 1947) comb. nov.; 289. A. subimpressus (Beier, 1955); 290. A. triangularis (Ellingsen, 1912) comb. nov.; 291. Geogarypus deceptor sp. nov.; 292. G. flavus (Beier, 1947) stat. nov., where green indicates confirmed localities and red unconfirmed published data.
Geogarypus liomendontus sp. nov.
Known from only two localities, both in isolated gorges just outside the northern edge of the MPA centre of endemism (Fig. 293). Both gorges were predominantly arid and contained dense indigenous tree stands growing next to very steep rock faces.
Habitat & Biology: All specimens were collected from cool, dry and shaded leaf litter located either against the trunks of trees or occasionally from deposits of litter between groups of rocks. Elevation: 760–785 m.
Geogarypus modjadji sp. nov.
Currently only known from its type locality (Fig. 294). Extensive sampling at other locations near the type locality failed to yield any further specimens. The locality is unique in that it contains the world’s largest concentration of a single species of cycad, known as the Modjadi palm (Encephalartos transvenosus). The area consists of a 305 ha reserve situated within a gorge, with a central stream that is mostly shaded by an isolated forest made up of cycads, up to 13 m in height (Grobbelaar, Meyer & Burchmore 1988). Due to the unique nature and isolation of this habitat it is believed that G. modjadi sp. nov. constitutes a short-range endemic species, restricted to this distinct forest.
Habitat & Biology: Moist leaf litter in undergrowth of cycad forest, particularly deposits among rock formations. Specimens were collected during the month of November. Elevation: 900 m.
Geogarypus octoramosus sp. nov.
Found primarily in the northern reaches of KwaZulu-Natal, the distribution of the species falls mostly within the MPA centre of endemism (Fig. 295). Although most of the locality data is situated either in the Grassland or Savanna biomes, most specimens were collected from leaf litter under bush clumps located either on top of rocky hills or next to rocky outcrops at altitudes above 1000 m. Other specimens were collected from Eucalyptus plantations or endemic Maputaland Sand Forest (Licuati).
Habitat & Biology: Cool and semi-moist shaded leaf litter, especially under thickets growing between rocky outcrops that result in leaf litter collecting in the crevices. Specimens have been recorded from January to May. Elevation: 100–1900 m.
Geogarypus olivaceus (Tullgren, 1907)
One of the few South African geogarypid species with substantial records from the interior of the country, G. olivaceus can be found in five provinces, with a distribution stretching as far south as the coast of the Western Cape and as far north as the Kruger National Park in Mpumalanga (Fig. 296). Specimens can easily be found within the grassland biome of central South Africa, particularly in the leaf litter of the African Wild Olive tree (Olea europaea subsp. africana) that commonly grows along rocky hills in the Free State at altitudes above 1100 m.
Habitat & Biology: Records show that the species is present throughout the year, with most being collected from cool, shaded and semi-moist to dry leaf litter in the central Grassland Biome of South Africa. Elevation: 20–1400 m.
Geogarypus tectomaculatus sp. nov.
Another potential short-range endemic, G. tectomaculatus sp. nov. seems to be confined to rocky outcrops along the edges of ridges in the highlands of Mpumalanga (Fig. 297). These particular rock formations form many crevices and shaded areas that allow shrubs to grow and deposit deep layers of leaf litter in an otherwise open, grass-dominated biome. It is within such deposits that this new species was most often found.
Habitat & Biology: Most often found within cool, dry and shaded leaf litter deposited in crevices around rock formations in open grassland. Specimens were also found in moist leaf litter of scattered indigenous forest patches. Collection records range from September to October. Elevation: 1200–1700 m.
Geogarypus variaspinosus sp. nov.
This is the only species found entirely within the Northern Cape (Fig. 298). It inhabits the leaf litter of shrubs on rocky hills and along rivers of both the Nama-Karoo and the adjacent savanna biomes of the province. The species seems to be more xerophilic than the other South African Geogarypidae, in that it is often found in very dry and sandy leaf litter.
Habitat & Biology: Other than its occurrence in very dry leaf litter in the arid Northern Cape, little is known about the biology of this new species. All collection records are for the winter month of June. Elevation: 900–1300 m.
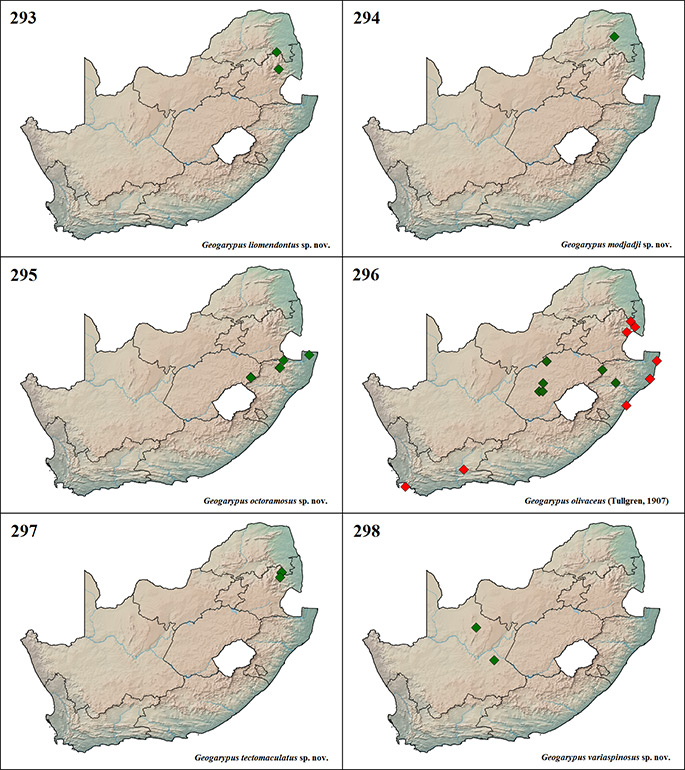
Figures 293–298. Geographical distribution of: 293. Geogarypus liomendontus sp. nov.; 294. G. modjadji sp. nov.; 295. G. octoramosus sp. nov.; 296. G. olivaceus (Tullgren, 1907); 297. G. tectomaculatus sp. nov.; 298. G. variaspinosus sp. nov., where green indicates confirmed localities and red unconfirmed published data.
South African Geogarypidae phylogenetics
The phylogeny of the Pseudoscorpiones has been a hotly debated topic amongst experts in the field since the late 19th century (Wheeler & Hayashi 1998). Historically it has been placed as a sister group to the Solifugae in the clade Haplocnemata. However, the internal phylogeny of the Pseudoscorpiones has not received much attention, with most early studies focussing on the non-numerical analysis of morphological features (Murienne et al. 2008).
The systematics of the group was modernised by the seminal works of Chamberlin (1929, 1930, 1931), who divided the Pseudoscorpiones into three suborders namely, Monosphyronida, Diplosphyronida and Heterosphyronida. Beier (1932) adopted Chamberlin’s scheme and proposed the equivalent suborders of Cheliferinea, Neobisiinea and Chthoniinea. These classification systems remained the norms in America and Europe, respectively, until the first cladistic analysis of the group was done by Harvey (1992). Using 126 characters he recognised 24 families. Furthermore, he placed these families within two suborders, based on either the presence (Iocheirata) or lack (Epiocheirata) of a venom apparatus on the chelal fingers.
With the advent of modern molecular techniques, the phylogeny of the group was once again subjected to scrutiny. Although molecular work was done on single species by authors such as Wilcox et al. (1997) and Wheeler & Hayashi (1998), phylogenetic analysis of pseudoscorpion species groups was conducted only recently (Zeh, Zeh & Bonilla 2003; Moulds et al. 2007). The most significant study is that done by Murienne et al. (2008), where through the use of two nuclear and one mitochondrial gene, the phylogenetic relationships of the major pseudoscorpion clades were investigated. Many of the superfamilies proposed in Harvey’s (1992) morphological cladistic analysis were found to be monophyletic, although the superfamilies Neobisiodea, Garypoidea and Cheliferoidea were not. Their study thus supported an observation made by Mallat & Giribet (2006), who found that although phylogenetic relationships derived from gene-sequences generally agree with traditional morphologically-derived relationships, the former can sometimes show startling differences, necessitating revisions of species’ morphology to account for their genetic relationships. The same discrepancy was noted in this study, as can be seen by the multiple new combinations, status updates and new species discovered after molecular phylogenetic analysis.
Both 18S rRNA and 28S rRNA have been extensively used throughout the years to infer phylogenetic relationships among arthropods (Wheeler & Hayashi 1998; Mallat & Giribet 2006), and recently among pseudoscorpions as well (Murienne et al. 2008). In this study 18S rRNA was discarded, both due to the potential for weak resolution at species level branches (Steiner & Müller 1996), and the costs involved. Only COI and 28S genes were used as they better reflect species level relationships.
Cytochrome C Oxidase Subunit I (COI) gene is one of the most popular genes currently used to infer phylogenetic relationships, and has been used by many authors in recent years in phylogeographic studies of pseudoscorpion populations (Wilcox et al. 1997; Zeh et al. 2003; Moulds et al. 2007; Murienne et al. 2008; Pfeiler et al. 2009; Van Heerden et al. 2013).
Amplification results
Of the 18 morphospecies sequenced, 17 yielded 28S rRNA PCR products and 17 yielded COI PCR products. After multiple PCR parameter adjustments, as well as genomic DNA extractions, we were unable to obtain 28S rRNA products for Afrogarypus minutus (Tullgren, 1907) comb. nov. or COI products for A. megamolaris sp. nov., resulting in their absence from the corresponding trees (Figs 299 & 300). Since both of the abovementioned species yielded results for at least one gene, they could still be incorporated within the concatenated tree (Fig. 301), and showed the same placement as other species with similar morphology. Positive PCR products of the 28 rRNA gene yielded sequences with post-trim and assembly lengths of 998–1077 bp, while the COI products yielded sequences 674–682 bp in length. All sequences were deposited in GenBank under the accession numbers shown in Table 1.
28S rRNA results
Phylogenetic analysis of the 28S rRNA gene sequences of the South African Geogarypidae using multiple alignment and tree construction models yielded trees that differed only slightly in topology, with the tree constructed using Geneious alignment and Neighbour Joining being chosen as the most congruent (Fig. 299).
Table 1. GenBank accession numbers for the nuclear 28S rRNA and mitochondrial COI sequences obtained during the study. Abbreviations: NBG – National Botanical Gardens; NP – National Park; NR – Nature Reserve.
| Species | GenBank Accession Numbers | Specimen Locality | ||
| 28S rRNA | COI | |||
| Afrogarypus (Beier, 1931) | ||||
| A. carmenae sp. nov. | KP297847 | KP331813 | Clanwilliam | |
| A. castigatus sp. nov. | KP297858 | KP331824 | St. Lucia | |
| A. excelsus (Beier, 1964) stat. nov. | KP297844 | KP331810 | Fort Fordyce NR | |
| A. impressus (Tullgren, 1907) | KP297845 | KP331811 | Tsitsikamma NP | |
| A. megamolaris sp. nov. | KP297854 | – | Makhado | |
| A. minutus (Tullgren, 1907) comb. nov. | – | KP331815 | Brenton-on-Sea | |
| A. purcelli (Ellingsen, 1912) comb. nov. | KP297850 | KP331817 | De Hoop NR | |
| A. robustus (Beier, 1947) comb. nov. | KP297851 | KP331818 | Kirstenbosch NBG | |
| A. subimpressus (Beier, 1955) | KP297846 | KP331812 | Cape Point NR | |
| A. triangularis (Ellingsen, 1912) comb. nov. | KP297852 | KP331819 | King William’s Town | |
| Geogarypus Chamberlin, 1930 | ||||
| G. deceptor sp. nov. | KP297853 | KP331820 | St. Lucia | |
| G. flavus Beier, 1947 stat. nov. | KP297848 | KP331814 | Jubilee Creek NR | |
| G. liomendontus sp. nov. | KP297843 | KP331809 | Abel Erasmus Pass | |
| G. modjadji sp. nov. | KP297855 | KP331821 | Modjadjikloof NR | |
| G. octoramosus sp. nov. | KP297859 | KP331825 | Vryheid | |
| G. olivaceus (Tullgren, 1907) | KP297849 | KP331816 | Bothaville | |
| G. tectomaculatus sp. nov. | KP297857 | KP331823 | Graskop | |
| G. variaspinosus sp. nov. | KP297856 | KP331822 | Douglas | |
The resulting 28S rRNA tree shows the presence of four major clades within the South African Geogarypidae, two clades corresponding with Afrogarypus and two clades corresponding to Geogarypus. The most distinctive feature of the tree is the inclusion of G. minutus, G. purcelli, G. robustus and G. triangularis in the Afrogarypus clade. The two subspecies, A. excelsus excellens and A. excelsus excelsus, are shown to be genetically identical. Geogarypus flavus, originally described as a separate species but later synonymised with G. olivaceus by Beier (1955), is once again resolved as a separate species, with the two species falling into different Geogarypus clades.
The tree also supported the presence of nine new species, with A. carmenae sp. nov., A. castigatus sp. nov. and A. megamolaris sp. nov. representing three new species of Afrogarypus, and G. deceptor sp. nov., G. liomendontus sp. nov., G. modjadji sp. nov., G. octoramosus sp. nov., G. tectomaculatus sp. nov. and G. variaspinosus sp. nov. placed in Geogarypus.

Figure 299. Neighbour Joining tree with 1000 Bootstrap repetitions of 28S rRNA sequences depicting phylogenetic relationships of South African Geogarypidae.
Morphologically, species forming the four major clades share the following similarities:
i – The species comprising the first Afrogarypus clade, consisting of A. excelsus excellens, A. excelsus excelsus, A. carmenae sp. nov., A. purcelli comb. nov., A. impressus and A. robustus comb. nov., all have chela with only slightly convex interior hand surfaces, chelal fingers with many small teeth, as well as either a well developed, deep dorsal sulcus, or concave dorsal region on the chela hand.
ii – The species in the second Afrogarypus clade, consisting of A. megamolaris sp. nov., A. triangularis comb. nov., A. subimpressus and A. castigatus sp. nov., have chela where the interior surface of the hand is enlarged to a globular strongly convex shape, with the exception of A. megamolaris sp. nov. They furthermore contain chelal fingers with fewer but more prominent teeth, and the presence of either a broader, shallower dorsal sulcus or a concave dorsal region. Afrogarypus triangularis comb. nov. and A. castigatus sp. nov. also show the presence of lateral abdominal sclerites in the males.
iii – The species comprising the first Geogarypus clade, consisting of G. olivaceus, G. octoramosus sp. nov. and G. variaspinosus sp. nov. have carapaces and tergites that are uniformly brown, with slightly lighter markings in only a small number of specimens. Males of the species all possess simple galea with no spinules, while the females of G. octoramosus sp. nov. and G. variaspinosus sp. nov. have galea with fewer than nine rami.
iv – In the second Geogarypus clade, G. flavus stat. nov. and G. tectomaculatus sp. nov. have tergites with distinct cream-coloured patches, female galea that are very broad, and male galea that possess spinules. Geogarypus deceptor sp. nov., G. liomendontus sp. nov. and G. modjadji sp. nov. furthermore have distinct cream patches on the posterior half of their carapaces, with all females in this group possessing galeae with nine rami. Geogarypus deceptor sp. nov. and G. modjadji males also possess galeae with rami.
COI results
Using the same parameters as the 28S rRNA analysis, the COI gene sequences of the South African Geogarypidae once again yielded trees that differed only slightly in topology, with the tree constructed using Geneious alignment and Neighbour Joining again being chosen as the most congruent (Fig. 300).
The COI tree differs from the 28S rRNA tree with regard to the topographical placement of the new species in particular, but supports many of the main results. These include the placement of G. minutus, G. purcelli, G. robustus and G. triangularis in the Afrogarypus clade; A. excelsus excellens and A. excelsus excelsus being genetically identical; and the re-validation of G. flavus stat. nov. as a distinct species. The nine new species are still distinct, and with the exception of A. castigatus sp. nov., group within the same genus clades as with the 28S rRNA results.
Of particular note is the placement of A. castigatus sp. nov. as a sister group to all the other South African Geogarypidae. Given the species’ unusual morphological adaptations, this placement is not unexpected. One troubling aspect of the tree is the very low Bootstrap values at some of the nodes, particularly between the taxa of the Geogarypus clade. Saturation of the COI gene is a possible explanation, as the mitochondrial genome evolves much faster than the nuclear genome (Burger, Gray & Lang 2003).
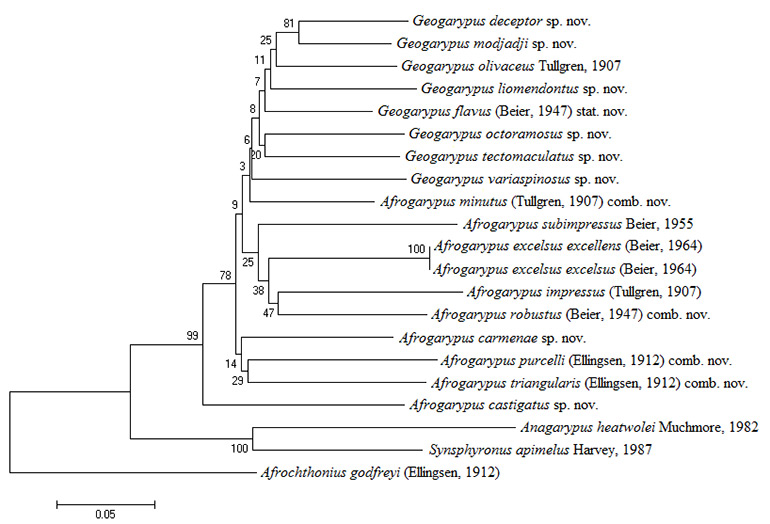
Figure 300. Neighbour Joining tree with 1000 Bootstrap repetitions of COI sequences depicting phylogenetic relationships of South African Geogarypidae.
Concatenated results
Super-gene alignments are one of two fundamentally different techniques used to construct phylogenetic trees from multiple genes, the other being the use of consensus trees. Gadagkar, Rosenberg & Kumar (2005) found that a combination of concatenation and Neighbour Joining analysis consistently yielded more accurate trees than consensus analysis. It was thus decided to follow the same technique and construct a tree from concatenated sequences of both the 28S rRNA and the COI genes to increase the accuracy of the phylogenetic analysis. After concatenation, the new super-genes were aligned using Geneious and the first consensus tree was constructed using Neighbour Joining with 1000 Bootstrap replicates.
The resulting tree (Fig. 301) shows that the Geogarypidae forms a monophylic clade separate to the Garypidae, and yielded four distinct clades, one corresponding to Geogarypus and three corresponding to Afrogarypus. The major trends of the separate 28S and COI trees are once again present and resulted in the following changes and additions:
i – Afrogarypus minutus comb. nov., A. purcelli comb. nov., A. robustus comb. nov. and A. triangularis comb. nov. are transferred to Afrogarypus. These combinations are supported by the presence of a concave area on the dorsal surface of the chela, present only within these species and A. castigatus sp. nov.
ii – Afrogarypus excelsus excellens and A. excelsus excelsus are synonymised under Afrogarypus excelsus stat. nov., due to the lack of any significant branching. The slight morphological differences used by Beier (1964) to distinguish between the two subspecies can best be attributed to intraspecific variation within the species. A well documented case supporting intraspecific variation was presented by Zeh & Zeh (1992) on the intra-brood variability in the chelal morphology of male Semeiochernes armiger (Chernetidae) (Balzan, 1892).
iii – Given both its phylogenetic separation and distinct morphological characters, G. flavus stat. nov. is removed from synonymy with G. olivaceus and re–validated as a distinct species.
iv – Despite its unique morphological adaptations and its phylogenetic separation, A. castigatus sp. nov. will be treated as a species of Afrogarypus, due to it possessing a concave dorsal area on the chela, consistent with that of other species of the genus. Its morphological synapomorphies, i.e. the presence of monotarsate leg pairs I and II, and a reduced trichobothrial compliment due to the absence of trichobothria isb, makes this species unique among South African pseudoscorpions, and this can possibly be explained as a simple case of paedomorphosis in a small species as both of the morphological reductions have been recorded in, and seem commonplace, in Garypidae (Harvey 1987a).
v – The following nine new species are recognised as they are both phylogenetically and morphologically distinct: A. carmenae sp. nov., A. castigatus sp. nov.; A. megamolaris sp. nov., G. deceptor sp. nov., G. liomendontus sp. nov., G. modjadji sp. nov., G. octoramosus sp. nov., G. tectomaculatus sp. nov. and G. variaspinosus sp. nov.
The particularly low Bootstrap values at some of the nodes necessitated a careful study of morphological features to help validate the phylogenetic findings. Fortunately, low values were present between taxa with very distinct morphological differences, including historically described species. Murienne et al. (2008) recommended the incorporation of pseudoscorpion-specific genes, or the design of specific primers instead of using universal varieties, to more accurately delimit species.
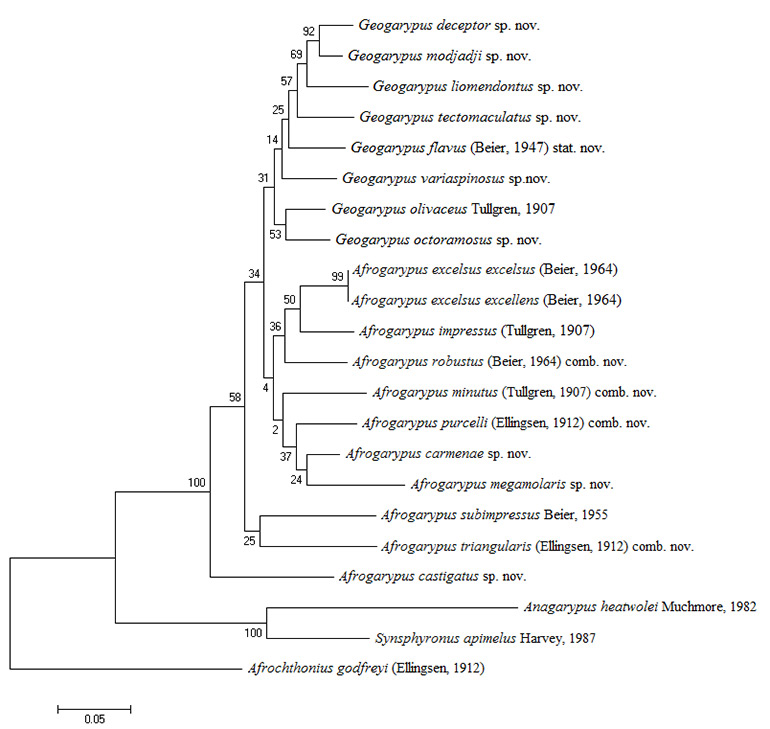
Figure 301. Neighbour Joining tree depicting phylogenetic relationships between taxa of South African Geogarypidae, constructed using concatenated sequences of both 28S rRNA and COI genes.
The results of the Bayesian analysis of the concatenated data (Fig. 302) correlates well with the previous trees, and is most similar to the 28rRNA consensus tree. It results in three major clades, one corresponding to Geogarypus and two corresponding to Afrogarypus. Both genera are again represented as paraphyletic and well defined by synapomorphic characters. Afrogarypus again includes A. minutus comb. nov., A. purcelli comb. nov., A. robustus comb. nov. and A. triangularis comb. nov., originally in Geogarypus, together with A. carmenae sp. nov., A. castigatus sp. nov. and A. megamolaris sp. nov., which represent new species. All the species share the presence of either a well developed dorsal sulcus or a concave dorsal depression. The rest of the species group within the Geogarypus clade and share chela that possess a convex dorsal surface. Of importance is the presence of two trichotomous splits within the tree, one within each clade, indicating that the concatenated sequences were inadequate to perfectly resolve all of the species. This again highlights the need for the use of new genes and loci to sufficiently resolve the phylogenetic relationships of pseudoscorpions.
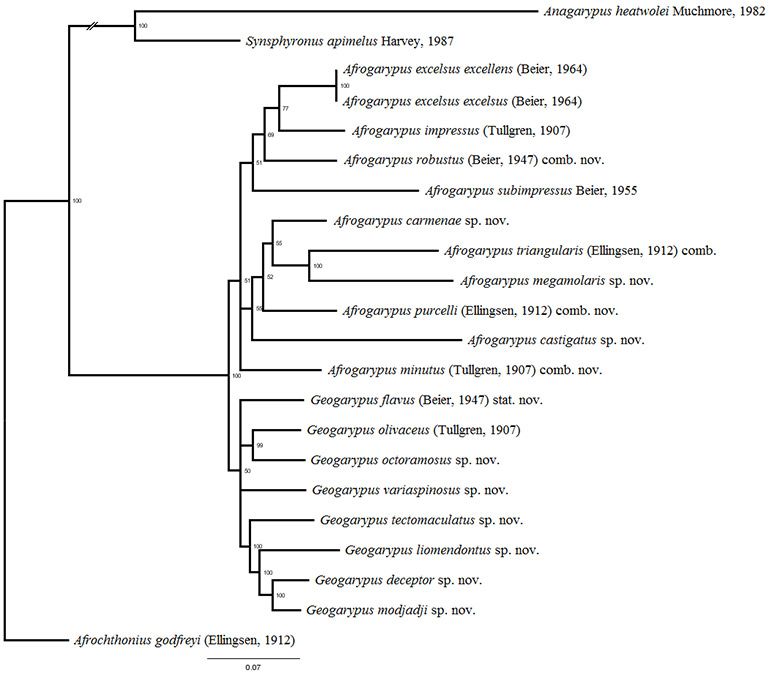
Figure 302. Bayesian analysis consensus tree depicting phylogenetic relationships between taxa of South African Geogarypidae, constructed using concatenated sequences of both 28S rRNA and COI genes.
Discussion
By incorporating a genetic section into this study, valuable insight was achieved with regards to the variability of morphological characters and their value in distinguishing between closely related species. Narrowing down the limits of these variations were major factors contributing to both the synonymisation of A. excelsus excellens, as well as the re-validation of G. flavus stat. nov. The above analysis furthermore resulted in the identification of cryptic species such as G. deceptor sp. nov., that on multiple occasions was mis-identified as either G. flavus stat. nov. or G. olivaceus in museum collections. Geogarypus modjadji sp. nov., another cryptic species that closely resembles G. liomendontus sp. nov., was also originally identified by its phylogenetic distinction. This led to the recognition of distinct differences in basal teeth morphology of their chelal fingers that can now be used to accurately distinguish the two species.
This study also presented the difficulty of finding and using distinct morphological characters in male Geogarypidae for identifying species, as the most distinct characters are found in females. Zeh & Zeh (1994) faced a similar situation in their study, where the uniformity of pseudoscorpion males masked the population’s genetic diversity. This issue can best be resolved using techniques to identify micro-morphological characters that aid in distinguishing between males (Muster, Schmarda & Blick 2004), as well as DNA barcoding.
In conclusion, the phylogenetic analysis of South African Geogarypidae highlighted, amongst other things, the hazards of using pure morphological techniques to infer relationships between pseudoscorpion species. When it comes to the systematics of the Pseudoscorpiones, previous studies showed that the sole use of either phylogenetic or morphological techniques lack the resolution to identify all complicated relationships among species (Zeh & Zeh 1992; Wilcox et al. 1997). Van Heerden et al. (2013) suggested that a combination of phylogenetic analysis and morphological analysis (using micro-morphological characters) should be used to infer relationships between pseudoscorpion species. The current study fully supports this holistic view for pseudoscorpion systematics.
Discussion
With the discovery of nine new species, bringing the total number of South African species to
18, South Africa is currently the most diverse country for Geogarypidae fauna. It is clear that large areas of central, northern and western South Africa require more intensive sampling, as few records are present from the Northern Cape, North West, Free State, Gauteng and Limpopo provinces. The use of canopy fogging proved invaluable for collecting specimens from habitat strata not accessible using more conventional sampling methods (leaf litter sifting, sweep netting, beating, pitfalls and hand collecting), yielding valuable insight into habitat flexibility in this family.
The most unique species discovered was A. castigatus sp. nov., the first South African geogarypid with both monotarsate front legs, as well as a reduced trichobothrial count. Interestingly, members of the Geogarypidae sharing these reduced features were already known, as Harvey (1986) described an Australian species (Geogarypus connatus) during his revision of the Geogarypidae that also possesses monotarsate leg pairs I and II, as well as the absence of tricobothria isb. Furthermore, all collection records of G. connatus were from under bark, indicating that this species may also be an obligate arboreal inhabitant, like A. castigatus sp. nov. Phylogenetic analysis of the South African and Australian geogarypid fauna will likely yield very interesting results.
New combinations of species into Afrogarypus necessitated an update of the diagnostic characters of the South African genera. Geogarypus is here defined by a uniformly convex dorsal chela hand surface and with a usual trichobothrial compliment of 8/4. Afrogarypus possess either a well developed sulcus or concave depression on the dorsal surface of the chelal hand, as well as the usual trichobothrial compliment of 8/4, with the exception of A. castigatus sp. nov., which presents distinct monotarsate anterior legs and the absence of trichobothria isb, resulting in a reduced trichobothrial compliment of 7/4.
The currently known distribution of Geogarypidae in South Africa correlates very closely with known centres of endemism, such as the Cape Floristic Region and Maputaland–Pondoland–Albany centres of endemism, and agrees with distribution patterns of other arachnids (Kuntner et al. 2008; Lotz 2009; De Bivort & Giribet 2010; Haddad & Wesołowska 2013; Wesołowska & Haddad 2013; Haddad 2014). Griswold (1991) eluded that the speciation seen today within South African Afromontane spiders could have resulted from fragmentation of once-persistent forest cover into isolated patches during periods of aridity, the most significant of these periods occurring during the Miocene-Pliocene and Pleistocene epochs. These isolation events would have had the same effects on the distribution and speciation of the Geogarypidae present at the time, and are most likely a major factor in explaining the distribution of modern species within isolated forest patches of the Maputaland-Pondoland-Albany centre of endemism along the eastern and southern parts of South Africa.
The genetic analysis in this study represents only the second phylogenetic analysis of any South African pseudoscorpions. The first study was recently published by Van Heerden et al. (2013) on members of the Olpiidae. Phylogenetic analysis of South African geogarypids followed many patterns observed by other authors working on similar pseudoscorpion groups, and yielded valuable information on the Geogarypidae as a whole. Most importantly, phylogenetic results corroborated the morphological analysis, and yielded the first holistic view on the systematics of the South African species of a family of pseudoscorpions. Although the phylogenetic analysis proved to be successful, difficulties with low Bootstrap values and the lack of gene amplification in Afrogarypus minutus (Tullgren, 1907) stat. nov. and A. megamolaris sp. nov. highlighted areas that need further research, including the identification of pseudoscorpion-specific genes and the development of group-specific primers.
This study furthermore highlighted the lack of pseudoscorpion expertise within South Africa, as could be seen by both the lack of even family-level identifications for many museum exemplars, as well as the many mis-identifications in South African collections. The difficulty of both tracking down and loaning type specimens was exacerbated by the apparent loss of the type specimens of A. impressus, A. purcelli comb. nov. and A. triangularis comb. nov. Literature indicated that the A. impressus type was housed together with the types for A. minutus and G. olivaceus, and although the last two types could be located within the Zoological Museum, Hamburg, the type of the first species was not present (M. Koch, pers. comm.). The same issue arose after a literature study suggested that the types for the Ellingsen species were housed within the Iziko South African Museum, Cape Town. After extensive searching only the types of A. robustus and G. flavus were located, but none of Ellingsen’s types. Harvey (1994) experienced a similar issue while searching for the type series of species in Chthoniidae and Tridenchthoniidae from Queensland, Australia, but was fortunate enough to ‘re-discover’ the lost types in the Department of Entomology of the University of Queensland.
In conclusion, this study aimed to provide the first holistic approach to the systematics of the South African species of a pseudoscorpion family, and thereby spearheads an endeavour to revise the entire fauna. The holistic approach furthermore yielded a more complete taxonomic view than either the morphological or phylogenetic analyses on their own. The discovery of nine new Geogarypidae species also suggests that similar results could be expected with the revisions of other pseudoscorpion families in South Africa. This study will act as a reference with regard to the techniques that can be used, possible difficulties that can be expected, as well as setting the basis for future technical improvements.
Acknowledgements
This study formed part of the M.Sc. of the first author at the University of the Free State, under supervision of the second author. The success of this project could only have been achieved through the contributions and effort of the following institutions and people: Dr Gaynor Dolman and Dr Mark S. Harvey of the Western Australian Museum, Perth (WAM), respectively for assistance with the molecular analysis and taxonomy of pseudoscorpions, and their hospitality during a visit to WAM in 2013; Dr Leon Lotz (National Museum, Bloemfontein) for his guidance and assistance during field work and in the loaning of exemplars; and Hanlie Grobler (Centre of Microscopy at the University of the Free State, Bloemfontein) for her help during S.E.M. work; the curators responsible for the loan of material that made this study possible, as well as all the curators from the institutions overseas that aided in tracking down holotypes. Special thanks to Carmen Neethling for her invaluable aid as a field assistant. This study was funded by a grant to the second author from the National Research Foundation of South Africa in the Thuthuka Programme (TTK2008050500003), including a study bursary to the first author, funds for field work, and international travel to attend the 19th International Congress of Arachnology in Taiwan and visit the Western Australian Museum in 2013.
References
BALZAN, L. 1892. Voyage de M. E. Simon au Venezuela (Decembre 1887-Avril 1888). Arachnides. Chernetes (Pseudoscorpiones). Annales de la Sociéte Entomologique de France 60: 497–552.
BEIER, M. 1931. Neue Pseudoscorpione der U. O. Neobisiinea. Mitteilungen aus dem Zoologischen Museum in Berlin 17: 299–318.
BEIER, M. 1932. Pseudoscorpionidea I. Subord. Chthoniinea et Neobisiinea. Tierreich 57: 1–258.
BEIER, M. 1947. Zur kenntnis der Pseudoscorpionidenfauna des Südlichen Afrika, inbesondere der Südwest- und Südafrikanischen trockengebiete. Eos 23: 285–339.
BEIER, M. 1955. Pseudoscorpionidea. In: Hanstrom, B., Brinck, P. & Rudebeck, G. (Eds.). South African Animal Life: Results of the Lund University Expedition in 1950–1951. Almquistand Wiksell, Stockholm. 263–328.
BEIER, M. 1957. Insects of Micronesia: Pseudoscorpionida. Third Edition. Burnise P. Bishop Museum, Honolulu, Hawaii. 64 pp.
BEIER, M. 1958. Pseudoscorpionidea (false scorpions) of Natal and Zululand. Annals of the Natal Museum 14: 155–187.
BEIER, M. 1963. Ordnung Pseudoscorpionidea (Afterskorpione). In: D’ Aguilar, J., Beier, M., Franz, I.H. & Raw, F. (Eds). Bestimmungsbücher zur bodenfauna Euripas I. Akademie-Verslag, Berlin. 313 pp.
BEIER, M. 1964. Weiteres zur kenntnis der Pseudoscorpioniden-Fauna des siidlichen Afrika. Annals of the Natal Museum 16: 30–90.
BEIER, M. 1966. Ergänzungen zur Pseudoscorpioniden-Fauna des südlichen Afrika. Annals of the Natal Museum 18: 455–470.
BERON, P. 2002. On the high altitude pseudoscorpions (Arachnida: Pseudoscorpionida) in the old world. Historia Naturalis Bulgarica 14: 29–44.
BURGER, G., GRAY, M.W. & LANG, B.F. 2003. Mitochondrial genomes: anything goes. Trends in Genetics 19: 709–716.
CHAMBERLIN, J.C. 1929. A synoptic classification of the false scorpions or chelaspinners, with a report on a cosmopolitan collection of the same. Part I. The Heterosphyronida (Chthoniidae) (Arachnida-Chelonethida). The Annals and Magazine of Natural History 4: 50–80.
CHAMBERLIN, J.C. 1930. A synoptic classification of the false scorpions or chela-spinners, with a report on a cosmopolitan collection of the same. Part II. The Diplosphyronida (Arachnida-Chelonethida). The Annals and Magazine of Natural History 10: 585–620.
CHAMBERLIN, J.C. 1931. The arachnid order Chelonethida. Stanford University Publications, Biological Sciences 7: 1–284.
COX, C.B. 2001. The biogeographic regions reconsidered. Journal of Biogeography 28: 511–523.
DE BIVORT, B.L. & GIRIBET, G. 2010. A systematic revision of the South African Pettalidae (Arachnida: Opiliones: Cyphophthalmi) based on a combined analysis of discrete and continuous morphological characters with the description of seven new species. Invertebrate Systematics 24: 371–406.
DEVASAHAYAM, S. & KOYA, K.M.A. 1994. Natural enemies of major insect pests of black pepper (Piper nigrum L.) in India. Journal of Spices & Aromatic Crops 3: 50–55.
DIPPENAAR-SCHOEMAN, A.S. & HARVEY, M.S. 2000. A check list of the pseudoscorpions of South Africa (Arachnida: Pseudoscorpiones). Koedoe 43: 89–102.
ELLINGSEN, E. 1904. On some pseudoscorpions from Patagonia collected by Dr Filippo Silvestri. Bollettino dei Musei di Zoologia ed Anatomia Comparata dell R. Università di Torino 19: 1–7.
ELLINGSEN, E. 1912. The pseudoscorpions of South Africa based on the collections of the South African Museum, Cape Town. Annals of the South African Museum 10: 75–128.
FOLMER, O., BLACK, M., HOEH, W., LUTZ, R. & VRIJENHOEK, R.C. 1994. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Molecular Marine Biology and Biotechnology 3: 294–299.
GADAGKAR, S.R., ROSENBERG, M.S. & KUMAR, S. 2005. Inferring species phylogenetics from multiple genes: concatenated sequence tree versus consensus gene tree. Journal of Experimental Zoology Part B: Molecular and Developmental Evolution 304B: 64–74.
GRISWOLD, C.E. 1991. Cladistic biogeography of Afromontane spiders. Australian Systematic Botany 4: 73–89.
GROBBELAAR, N., MEYER, J.J.M. & BURCHMORE, J. 1988. Coning and sex ration of Encephalartos transvenosus at the Modjadji Nature Reserve. South African Journal of Botany 55: 79–82.
HADDAD, C.R. 2014. A revision of the endemic South African sac spider genus Lessertina Lawrence, 1942 (Araneae: Eutichuridae). Zootaxa 3873: 82–92.
HADDAD, C.R. & DIPPENAAR-SCHOEMAN, A.S. 2009. A checklist of the non-acarine arachnids (Chelicerata: Arachnida) of the De Hoop Nature Reserve, Western Cape Province, South Africa. Koedoe 51: 1–9.
HADDAD, C.R. & DIPPENAAR-SCHOEMAN, A.S. 2015. Diversity of non-acarine arachnids of the Ophathe Game Reserve, South Africa: testing a rapid sampling protocol. Koedoe 57: 1–15.
HADDAD, C.R., DIPPENAAR-SCHOEMAN, A.S. & WESOŁOWSKA, W. 2006. A checklist of the non-acarine arachnids (Chelicerata: Arachnida) of the Ndumo Game Reserve, Maputaland, South Africa. Koedoe 49:1–22.
HADDAD, C.R. & WESOŁOWSKA, W. 2013. Additions to the jumping spider fauna of South Africa. Genus 24: 459–501.
HALL, B.G. 2013. Building phylogenetic trees from molecular data with MEGA. Molecular Biology and Evolution 30: 1229–1235.
HANSEN, H.J. 1893. Organs and characters in different orders of arachnids. Entomologiske
Meddelelser 4: 135–251.
HARVEY, M.S. 1981. Geogarypus rhantus sp. nov. (Pseudoscorpionida: Garypidae: Geogarypinae); a generic addition to the Australian fauna. Memoirs of the Queensland Museum 20: 279–283.
HARVEY, M.S. 1986. The Australian Geogarypidae, new status, with a review of the generic classification (Arachnida: Pseudoscorpiones). Australian Journal of Zoology 34: 753–778.
HARVEY, M.S. 1987a. A revision of the genus Synsphyronus Chamberlin (Garypidae, Pseudoscorpionida, Arachnida). Australian Journal of Zoology 126: 1–99.
HARVEY, M.S. 1987b. Redescriptions of Geogarypus bucculentus Beier and Geogarypus pustulatus Beier (Geogarypidae: Pseudoscorpionida). Bulletin of the British Arachnological Society 7: 137–144.
HARVEY, M.S. 1991. Catalogue of the Pseudoscorpionida. Manchester University Press, Manchester. 726 pp.
HARVEY, M.S. 1992. The phylogeny and classification of the Pseudoscorpionida (Chelicerata: Arachnida). Invertebrate Taxonomy 6: 1373–1435.
HARVEY, M.S. 1994. Rediscovered pseudoscorpion type material described by Beier from south-eastern Queensland (Arachnida: Pseudoscorpionida). Memoirs of the Queensland Museum 37: 155–156.
HARVEY, M.S. 1996. The biogeography of Gondwanan pseudoscorpions (Arachnida). Revue Suisse de Zoologie, hors série 1: 255–264.
HARVEY, M.S. 2000. From Siam to Rapa Nui – the identity and distribution of Geogarypus longidigitatus (Rainbow) (Pseudoscorpiones: Geogarypidae). Bulletin of the British Arachnological Society 11: 377–384.
HARVEY, M.S. 2010. Redescription of Geogarypus irrugatus from Sumatra (Pseudoscorpiones: Geogarypidae). Journal of Arachnology 38: 383–386.
HARVEY, M.S. 2013. Pseudoscorpions of the world, version 3.0. Western Australian Museum, Perth. (http://www.museum.wa.gov.au/catalogues/pseudoscorpions)
HARVEY, M.S., HUEY, J., HILLYER, M., MCINTYRE, E. & GIRIBET, G. 2016. The first
troglobitic species of Gymnobisiidae (Pseudoscorpiones, Neobisioidea), from Table Mountain (Western Cape Province, South Africa) and its phylogenetic position. Invertebrate Systematics 30: 75–85.
HENDERICKX, H. 2005. A new Geogarypus from Baltic amber (Pseudoscorpiones: Geogarypidae). Phegea 33: 87–92.
HENDERICKX, H. & PERKOVSKY, E.E. 2012. The first geogarypid (Pseudoscorpiones, Geogarypidae) in Rovno amber (Ukraine). Vestnik Zoologii 46: e33–e36.
HEWITT, J. & GODFREY, R. 1929. South African pseudoscorpions of the genus Chelifer. Annals of the Natal Museum 6: 305–336.
HU, J.F. & ZHANG, F. 2012. Description of two new Stenohya species from China (Pseudoscorpiones, Neobisiidae). ZooKeys 213: 79–91.
JUDSON, M.L.I. 2007. A new and endangered species of the pseudoscorpion genus Lagynochthonius from a cave in Vietnam, with notes on chelal morphology and the composition of the Tyrannochthoniini (Arachnida, Chelonethi, Chthoniidae). Zootaxa 1627: 53–68.
KOCH. L. 1873. Uebersichtliche Darstellung der europäischen Chernetiden (Pseudoscorpione). Bauer and Raspe, Nürnberg. 68 pp.
KUNTNER, M., HADDAD, C.R., ALJANČIČ, G. & BLEJEC, A. 2008. Ecology and web allometry of Clitaetra irenae, an arboricolous African orb-weaving spider (Araneae, Araneoidea, Nephilidae). Journal of Arachnology 36: 583–594.
LAWRENCE, R.F. 1935. A cavernicolous false scorpion from Table Mountain, Cape Town. Annals & Magazine of Natural History: Series 10 15: 549–555.
LAWRENCE, R.F. 1967. The pseudoscorpions (False-scorpions) of the Kruger National Park. Koedoe 10: 87–91.
LOTZ, L.N. 2009. Harvestmen (Arachnida: Opiliones) in southern Africa – an annotated catalogue with notes on distribution. Navorsinge van die Nasionale Museum, Bloemfontein 25: 1–46.
MAHNERT, V. 1988. Die pseudoskerpione (Arachnida) Kenyas. Familien Withiidae und Cheliferidae. Tropical Zoology 1: 39–89.
MAHNERT, V. 2011. A nature’s treasury: pseudoscorpion diversity of the Canary Islands, with the description of nine new species (Pseudoscorpiones, Chthoniidae, Cheiridiidae) and new records. Revista Ibérica de Aracnología 19: 27–45.
MAHNERT, V., DI IORIO, O., TURIENZO, P. & PORTA, A. 2011. Pseudoscorpions (Arachnida) from Argentina. New records of distributions and habitats, corrections and an identification key. Zootaxa 2881: 1–30.
MALLAT, J. & GIRIBET, G. 2006. Further use of nearly complete 28S and 18S rRNA genes to classify Ecdysozoa: 37 more arthropods and a kinorhynch. Molecular Phylogenetics and Evolution 40: 772–794.
MANSELL, M.W. & ERASMUS, F.N. 2002. Southern African biomes and the evolution of Palparini (Insecta: Neuroptera: Myrmeleontidae). Acta Zoologica Academiae Scientiarum Hungaricae 48: 175–184.
MILLER, S.A., DYKES, D.D. & POLESKY, H.F. 1988. A simple salting out procedure for extracting DNA from nucleated cells. Nucleic Acids Research 16: 1215.
MOULDS, T.A., MURPHY, N., ADAMS, M., REARDON, T., HARVEY, M.S., JENNINGS, J. & AUSTIN, D. 2007. Phylogeography of cave pseudoscorpions in southern Australia. Journal of Biogeography 34: 951–962.
MUCHMORE, W.B. 1982. Pseudoscorpionida. In: Parker, S.P. (Ed.). Synopsis and classification of living organisms. Second Edition. McGraw-Hill, New York, U.S.A. 96–102.
MUCINA, L. & RUTHERFORD, M.C. 2006. The vegetation of South Africa, Lesotho and Swaziland. Strelitzia 19. South African National Biodiversity Institute, Pretoria. 808 pp.
MURIENNE, J., HARVEY, M.S. & GIRIBET, G. 2008. First molecular phylogeny of the major clades of Pseudoscorpiones (Arthropoda: Chelicerata). Molecular Phylogenetics and Evolution 49: 170–184.
MURTHY, V.A. & ANANTHAKRISHAN, T.N. 1977. Indian Chelonethi. Oriental Insects Monograph 4: 1–183.
MUSTER, C., SCHMARDA, T. & BLICK, T. 2004. Vicariance in a cryptic species pair of European pseudoscorpions (Arachnida, Pseudoscorpiones, Chthoniidae). Zoologischer Anzeiger 242: 299–311.
PERERA, S.J., RATNAYAKE-PERERA, D. & PROCHEŞ, Ş. 2011. Vertebrate distributions indicate a greater Maputaland–Pondoland–Albany region of endemism. South African Journal of Science 107: 1–15.
PETROV, B.P. 1997. A review of Bulgarian pseudoscorpions (Arachnida, Pseudoscorpionida). In: Zabka, M. (Ed.). Procedings of the 16th European Colloquium of Arachnology. Wyźsza Szkola Rolnicko-Pedagogniczna, Siedlce. Pp 261–269.
PFEILER, E., BITLER, B.G., CASTREZANA, S., MATZKIN, L.M. & MARKOW, T.A. 2009. Genetic diversification and demographic history of the cactophilic pseudoscorpion Dinocheirus arizonensis from the Sonoran Desert. Molecular Phylogenetics and Evolution 52: 133–141.
PICKER, M.D. & SAMWAYS, M.J. 1996. Faunal diversity and endemicity of the Cape Peninsula, South Africa – a first assessment. Biodiversity and Conservation 5: 591–606.
ROEWER, C.F. 1936. Chelonethi order Pseudoskorpione. In: Bronns, H.G. (Ed.). Dr. H. G. Bronn’s Klassen und ordnungen des Tierreichs. Akademische Verlagsgesellschaft, Leipzig. Pp. 1–160.
ROEWER, C.F. 1937. Chelonethi order Pseudoskorpione. In: Bronns, H.G. (Ed). Dr. H. G. Bronn’s Klassen und ordnungen des Tierreichs. Akademische Verlagsgesellschaft, Leipzig. Pp. 161–320.
SIMON, E. 1879. Les Arachnides de France VII: Les Ordres des Chernetes, Scorpiones et Opiliones. Librairie Encyclopédique de Roret, Paris. 322 pp.
SPAULL, V.W. 1979. Distribution of soil arthropods on Aldabra Atoll. Philosophical Transactions of the Royal Society of London (B) 285: 109–117.
ŠŤÁHLAVSKÝ, F., KRÁL, J., HARVEY, M.S. & HADDAD, C.R. 2006. A karyotype study on the pseudoscorpion families Geogarypidae, Garypinidae and Olpiidae (Arachnida: Pseudoscorpiones). European Journal of Entomology 103: 277–289.
ŠŤÁHLAVSKÝ, F., KRÁL, J., HARVEY, M.S. & HADDAD, C.R. 2012. The first cytogenetic characterisation of atemnids: pseudoscorpions with the highest chromosome numbers (Arachnida: Pseudoscorpiones). Cytogenetic and Genome Research 137: 22–30.
STEINER, G. & MÜLLER, M. 1996. What can 18S rDNA do for Bivalve phylogeny? Journal of Molecular Evolution 43: 58–70.
TÖMÖSVÁRY, O. 1882. A magyar fauna álskorpiói (Pseudoscorpiones faunae hungaricae). Magyar Tudományos Akadémia Matematikai és Természettudományi Közlemények 18: 135–256.
TULLGREN, A. 1907a. Zur kenntnis aussereuropäischer Chelonethiden des Naturhistorischen Museums in Hamburg. Mitteilungen aus dem Naturhistorischen Museum in Hamburg 24: 21–75.
TULLGREN, A. 1907b. Chelonethiden aus Natal und Zululand. In: Wirén, A. (Ed.). Zoologiska studier tillagnade Professor T. Tullberg. Almquist and Wiksells, Uppsala. Pp. 216–236.
TURIENZO, P., DI IORIO, O. & MAHNERT, V. 2010. Global checklist of pseudoscorpions (Arachnida) found in birds’ nests. Revue suisse de Zoologie 117: 557–598.
VAN HEERDEN, J., TAYLOR, P.J. & VAN HEERDEN, C. 2013. Genetic differentiation in Horus Chamberlin (Arachnida: Pseudoscorpiones: Olpiidae) as indicated by mitochondrial DNA analysis. African Zoology 48: 351–358.
WEIDNER, H. 1959. Die Entomologischen Sammlungen des Zoologischen Staatsinstituts und Zoologischen Museums Hamburg. I. Teil. Pararthropoda und Chelicerata I. Mitteilungen aus dem Hamburgischen Zoologischen Museum und Institute 57: 89–142.
WESOŁOWSKA, W. & HADDAD, C.R. 2013. New data on the jumping spiders of South Africa (Araneae: Salticidae). African Invertebrates 54: 177–240.
WHEELER, W.C. & HAYASHI, C.Y. 1998. The phylogeny of the extant chelicerate orders. Cladistics 14: 173–192.
WILCOX, T.P., HUGG, L., ZEH, J.A. & ZEH, D.W. 1997. Mitochondrial DNA sequencing reveals extreme genetic differentiation in a cryptic species complex of neotropical pseudoscorpions. Molecular Phylogenetics and Evolution 7: 208–216.
WITH, C.J. 1906. The Danish expedition to Siam 1899-1900. 111. Chelonethi. An account of the Indian false-scorpions together with studies on the anatomy and classification of the order. Oversigt over det Konigelige Danske Videnskabernes Selskabs Forhandlinger 3: 1–214.
ZEH, J.A. & ZEH, D.W. 1992. Are sexually-selected traits reliable species characters? Implications of intra-brood variability in Semeiochernes armiger (Balzan) (Pseudoscorpionida: Chernetidae). Bulletin of the British Arachnological Society 9: 61–64.
ZEH, J.A., ZEH, D.W. & BONILLA, M.M. 2003. Phylogeography of the harlequin beetle-riding pseudoscorpion and the rise of the Isthmus of Panamá. Molecular Ecology 12: 2759–2769.
ZEH, D.W. & ZEH, J.A. 1994. When morphology misleads: Interpopulation uniformity in sexual selection masks genetic divergence in harlequin beetle-riding pseudoscorpion populations. Evolution 48: 1168–1182.



Comments are closed.