
Dawid H. de Swardt1, Department of Ornithology, National Museum, Bloemfontein; Alan Lee2, DST/NRF Centre of Excellence at the Percy FitzPatrick Institute of African Ornithology, University of Cape Town; Hennie J.B. Butler3, Department of Zoology & Entomology, University of the Free State, Bloemfontein; H. Dieter Oschadleus4, School of Life Sciences, University of KwaZulu-Natal, Pietermaritzburg, South Africa
Abstract
Karoo Prinia (Prinia maculosa) and Drakensberg Prinia (Prinia hypoxantha) from various localities in South Africa and Lesotho were studied. Biometric data were obtained mainly during ringing studies. Karoo Prinia biometrics were analysed at subspecies level (P. m. maculosa, P. m. exultans, P. m. psammophila). Prinia m. maculosa males were relatively longer-winged, had longer central tail feathers (T1), longer total head length, and greater mean body mass than females. Prinia m. exultans male wing and central tail feathers (T1) were relatively longer than females. Both sexes of Prinia m. exultans were also found to be relatively longer-winged, have longer central tail feathers (T1) and greater body mass than P. m. maculosa. Prinia hypoxantha males were larger than females in all biometric parameters, including body mass. The biometric data obtained will be useful for sexing and aging birds during ringing studies. Support for the separate species status of P. hypoxantha (in north-eastern Free State, Mpumalanga and KwaZulu-Natal) is indicated by the fact that it tended to be longer-winged and longer-billed than the parapatric P. m. exultans (eastern Free State and Lesotho). An analysis of stomach contents of P. m. exultans and P. hypoxantha indicated that several insect orders were represented in the diet of both species. Coleopterans (beetles, in 78% of stomachs) and hemipterans (true bugs, 20%) were the most frequently recorded prey items for P. m. exultans, and coleopterans (32%) and hymenopterans (especially ants) (18%) were the most important for P. hypoxantha. Spiders were also eaten by both species and were present in as many as 36% of P. hypoxantha stomachs. Plant seeds were present in the stomachs of 13% of P. m. exultans and 36% of P. hypoxantha.
INTRODUCTION
The two species of ‘spotted’ prinias, Karoo Prinia Prinia maculosa (Boddaert) and Drakensberg Prinia Prinia hypoxantha (Sharpe) were initially described as a single species with four subspecies. Clancey (1989) reviewed the complex and found that the north-eastern form was a well-characterised allospecies (P. hypoxantha) distinct from three subspecies of P. maculata (P. m. maculosa, P. m. psammophila Clancey, 1989, P. m. exultans Clancey, 1989) which he recognised on the basis of three criteria, namely upper-part colouration, breast streaking, and absence or presence of a grey wash to the flanks. This classification was followed by Urban, Fry & Keith (1997) and Dean (2005a,b).
Prinia m. maculosa occurs in the Western, Northern and Eastern Cape provinces; P. m. psammophila occurs from southern Namibia to the western parts of the Northern Cape (Chittenden, Allan & Weiersbye 2012; Dean 2005a); and P. m. exultans occurs mainly in karoo areas of the Northern Cape, southern and eastern parts of the Free State, as well as the escarpment areas of Lesotho, Eastern Cape and KwaZulu-Natal where its range overlaps with P. hypoxantha (Chittenden, Allan & Weiersbye 2012; De Swardt 2010; DDS unpubl. data). The Drakensberg Prinia inhabits rank grass, bracken shrub and forest edges in high altitude areas of the Grassland Biome in Lesotho and the South African provinces of KwaZulu-Natal, Free State, Mpumalanga and Limpopo (Clancey 1997; Dean 2005a,b). Possible hybridisation between these two species occurs in Sani Pass and Garden Castle Caves Nature Reserve on the Drakensberg escarpment (Dean 2005b; De Swardt 2010; DDS unpubl data).
Morphometric data and information on the diet of these two species are limited. Previously published biometric data for Karoo Prinia are available for a sample of only 20 males and 16 females from the Western Cape (P. m. maculosa), including mass data for only nine males and four females (Dean 2005a; Urban, Fry & Keith 1997). No published biometric data are available for the P. m. exultans group in the Free State and Lesotho, or P. m. psammophila (Dean 2005a). Biometric data for Drakensberg Prinia are restricted to a small sample of unsexed birds, with mass data available for only eight males and four females (Dean 2005b). Our study presents a larger sample of biometric and body mass data for both the Karoo Prinia (mostly Free State and Western Cape areas) and Drakensberg Prinia. This data will be a useful guide for the research community and citizen scientists for aging and sexing during ringing and other studies. The biometrics of Karoo Prinia (P. m. exultans) of the eastern Free State and adjacent areas represents new information for this taxon, allowing for a comparison with related Prinia taxa.
Birds of colder and higher altitude areas are generally larger and heavier than those of the same species at warmer and lower altitudes, a phenomenon referred to as Bergmann’s Rule (Blackburn, Gaston & Loder 1999). This was also investigated in the present study.
While foraging behaviour of both species has been described, there is limited data available based on stomach contents (Dean 2005a,b; Kopij 2005). The present study investigates the diet of these two prinia species based on the stomach contents of a larger sample of birds, mostly from the Free State and KwaZulu-Natal.

MATERIALS AND METHODS
Study areas.–Karoo and Drakensberg Prinias were captured with mist nets during bird ringing studies (DDS, AL & HDO) at various localities in Lesotho and the South African provinces of Free State, KwaZulu-Natal, Western Cape and Mpumalanga, from October 1992 to March 2016 (for DDS data, see De Swardt 2010) (see Fig.1 for localities). Study sites in the Free State and KwaZulu-Natal were mainly Leucosidea sericea woodland and shrublands dominated by Olea and Buddleja species. Study sites in the Western Cape were located in fynbos vegetation on mountain slopes and drainage lines along streams.
Material examined.–Karoo Prinia biometrics were analysed at subspecies level. Prinia m. maculosa biometrics were obtained for the Western Cape during the period March 1999 to March 2016 during bird ringing studies in fynbos and wetland habitats (AL & HDO). Prinia m. exultans data are for the Eastern Cape, Free State, Lesotho and KwaZulu-Natal, obtained mainly from September 1991 to May 2015 (DDS). Small samples of P. m. psammophila biometrics were obtained from the curators of Ditsong National Museum of Natural History (Pretoria) and Durban Natural Science Museum, Durban (for Port Nolloth and Springbok, Northern Cape) and from SAFRING bird ringing data for the Northern Cape and southern Namibia (H. Kolberg). Prinia hypoxantha data are for the eastern Free State, KwaZulu-Natal and Mpumalanga, obtained from November 1990 to February 2016 (DDS & K. Raymakers).
In certain areas of the Free State, Karoo and Drakensberg Prinias were collected (with 4.10 combination rifle, or euthanised after capture in mist nets) as part of an ongoing project to obtain a representative collection of study skins for the Ornithology Department at the National Museum. The specimens were measured and weighed, and later prepared as study skins (or skeletons). Stomach contents of these specimens were preserved in 70% ethanol for future analysis. Birds were sexed during the skinning process, males by the presence of testes and females by the presence of ovaries. Skins already in the collection (collected from 1974 to 1994) were re-examined and measurements taken; mass data was available on the specimen tags. Specimens were measured upon collection from 1995 onwards.
Measurements and body mass.–The following standard measurements were recorded during ringing sessions and upon collection, as described by De Beer et al. (2000). Culmen length was measured from the skull-line to tip of the bill, and total head length measured from the back of the head to the tip of the bill, using vernier calipers. The tarsus was measured from the notch of the intertarsal joint to the lower edge of the last complete scale before the toes diverge. Wing length was measured from the shoulder of the closed wing to the longest primary feather, measured with a wing ruler, and both non-moulting shortest outer, and longest inner, tail lengths were measured. Body mass was recorded with a Pesola spring balance or an electronic scale (nearest 0.1 g).
Ringed birds were sexed based on wing and tail length criteria using Dean (2005a,b) as a guide, and also by using sexed study skins (gonads). Ringing data for P. m. maculosa were compared with those presented by Dean (2005a). Using measurements from sexed study skin specimens, birds with wing length >53.0 mm and tail length >70.0 mm were considered as males, and this was also used as a guide in sexing other prinias. Biometrics were investigated after measuring live birds and study skins. Shrinkage of preserved specimens was expected to be minimal.
Stomach content analysis.–A dietary analysis was performed on the stomach contents of 45 Prinia m. exultans (25 males, 15 females, three unsexed individuals and two juveniles) collected in Free State and Underberg in north-western KwaZulu-Natal from October 1998 to May 2015; and 22 P. hypoxantha (15 males, six females and one unsexed bird) collected in eastern Free State and the Sani Pass area of western KwaZulu-Natal from February 1999 to March 2012. Dietary items were sorted with the aid of a Zeiss stereoscopic dissecting microscope. Where possible, arthropods were identified to order and family level. The classification of insects followed the taxonomy of Scholtz & Holm (1985). All identified material was oven-dried at 75ºC in an Inc-O-Mat laboratory oven for 48 hr using a Metler P 160 NTop Loading Analytical Balance, and the dry mass of each arthropod order and family determined.
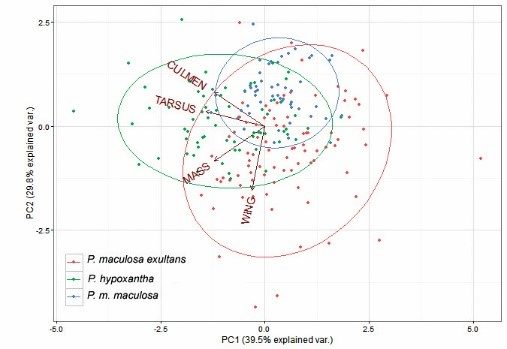
Statistical analysis.– Biometric data for P. hypoxantha and the two subspecies P. m. maculosa and P. m. exultans were all analysed separately. Limited data for a small sample of P. m. psammophila were not included in the multivariate analysis. A t-test, and t-test for independent samples by groups, between biometric data for P. maculosa subspecies and P. hypoxantha were performed using Statistica 13 (Dell Inc 2015). Pearsons’ correlation tests were used to examine the data of the two species and the subspecies of P. maculosa. Finally, we performed a principal components analysis (PCA) on biometric data (mass; wing, culmen and tarsus lengths) for P. m. maculosa, P. m. exultans and P. hypoxantha to explore the confidence of biometric data alone for identification.
RESULTS
BIOMETRICS
Karoo Prinia Prinia maculosa maculosa
Biometric data for 97 male and 159 female P. m. maculosa are presented in Table 1. Male mean wing lengths (x̄ = 53.8 mm) were significantly greater than those of females (x̄ = 51.3 mm) (t = 10.767; df = 253; P < 0.05). The longest tail feather (T1) of males (x̄ = 75.2 mm) was also significantly longer than that of females (x̄ = 66.6 mm; t = 10.467; df = 168; P < 0.05). Total head lengths between the sexes also differed significantly (t = 2.253; df = 127; P < 0.05), with longer heads in males. Males have a larger mean body mass (x̄ = 9.44) than females (x̄ = 9.18; t = 2.320; df = 200; P < 0.05). No significant differences exist between the sexes in culmen and shorter (T5) tail feather lengths; or between male wing and longest tail feather (T1) lengths (Table 1).
Karoo Prinia Prinia maculosa exultans
Biometric data for 89 male and 48 female P. m. exultans are presented in Table 2. Male wing (x̄ = 55.2 mm) and tail (T1) lengths (x̄ = 74.8 mm) were significantly greater than female wing (x̄ = 52.4 mm) and tail (x̄ = 67.5 mm) lengths (wing: t = 5.752; df = 135; P < 0.05; tail: t = 5.395; df = 132; P < 0.05). Culmen, total head and tarsus lengths, as well as body mass, did not differ significantly between the sexes.
Significant correlations between male wing and longest tail feather (T1) lengths were recorded (male: r = 0.214; females: r = 0.332) between P. m. exultans and P. hypoxantha (Tables 2 & 4).
Prinia m. exultans was relatively longer-winged, with higher mean body mass, than P. m. maculosa in both sexes (see Tables 1 & 2). A significant difference between the males of the two subspecies was evident in culmen and wing lengths, and mass (culmen: t = -3.852, df = 124, P < 0.05; wing: t = 2.881, df = 220, P < 0.05; mass: t = 5.081, df = 181, P < 0.05). In females, culmen and wing lengths and mass also differed significantly between the two subspecies (culmen: t = -5.595, df = 158, P < 0.05; wing; t = 3.851, df = 200, P < 0.05; mass: t = 5.758, df = 185, P < 0.05).


Karoo Prinia Prinia maculosa psammophila
Biometric data for 22 unsexed P. m. psammophila are presented in Table 3. This is based on museum specimens and ringing data (from SAFRING ringers). The sample sizes are considered too small for analysis.
Drakensberg Prinia Prinia hypoxantha
Biometric data for 72 male and 37 female P. hypoxantha is presented in Table 4. A significant difference in all biometrics (except tarsus length) between sexes was evident (t-test; all P < 0.05). Male wing (x̄ = 54.2 mm) and tail (T1) lengths (x̄ = 70.6 mm) were significantly greater than female wing (x̄ = 51.4 mm) and tail (x̄ = 62.0 mm) lengths (wing: t = 8.081; df =107; P < 0.05; tail: t = 5.37; df = 104; P < 0.05).
Biometric data for P. hypoxantha was also compared with P. m. exultans, since the geographical ranges of the two species overlap along the Eastern Cape and KwaZulu-Natal escarpments (Dean 2005a,b). All biometric parameters (culmen, total head, tarsus, wing and tail lengths) in males differed significantly between the two species (P < 0.05). No differences were found in male mass. Significant differences were noted in wing (t = 2.210, df = 93, P < 0.05) and tail (T1) (t = 2.885, df = 91, P < 0.05) lengths of males and females of the two species. In females, all biometric parameters (culmen, total head, tarsus, wing, tail lengths and mass) differed significantly between the two species (all P < 0.05). Prinia hypoxantha tended to be longer-billed with longer total head length in both sexes compared to P. m. exultans, although the latter tended to have longer tails in both sexes.

However, the PCA indicated that while males and females of the different species and subspecies form discernible clusters in ordination space (Fig. 2, Table 5a, b), there is considerable overlap in these metrics. Prinia hypoxantha was found to be significantly heavier than P. m. maculosa (lower altitude areas) in both sexes (males: t = 6.802, df = 169, P < 0.05; females: t = 12.072, df = 166, P < 0.05), which confirms Bergmann’s Rule. The t-test also confirms Bergmann’s Rule on species level between P. hypoxantha and P. maculosa (all regions) (males: t = -4.691, df = 222, P < 0.05; females: t = -7.759, df = 261, P < 0.05).
DIET
Diet composition (prey categories) is presented in Table 6. A total of nine arthropod orders were represented in the diet of P. m. exultans and six in the diet of P. hypoxantha. Although the diets of both species were dominated by insects, there were notable differences between the two taxa. Coleopterans (beetles, in 78% of stomachs) and hemipterans (true bugs, 20%) were the most frequently recorded prey items for P. m. exultans, and coleopterans (32%) and hymenopterans (especially ants) (18%) were the most frequent for P. hypoxantha. Spiders were also eaten by both species and were present in as many as 36% of P. hypoxantha stomachs. Plant matter in the form of seeds was also ingested by both prinia species, but in the case of P. hypoxantha this occurred much more frequently (36.3% versus 13.3%). Two P. m. exultans stomachs and one P. hypoxantha stomach also contained inorganic matter.
Coleopteran beetles formed the greatest proportion of total dietary mass for P. m. exultans (0.083 g; frequency 36.7%) of 0.234 g (Table 6). In terms of dietary mass, the most significant arthropod orders for P. m. exultans were Coleoptera, Hymenoptera and Hemiptera, and for P. hypoxantha they were Hymenoptera and Arachnida. In terms of frequency for P. hypoxantha, less coleopterans and more Araneae and Hymenoptera were recorded from stomachs.



prinia (P. m. maculosa, P. m. exultans, P. hypoxantha).
DISCUSSION
Prinia m. exultans, which occurs mainly on the escarpment of the eastern Free State, KwaZulu-Natal and Lesotho, was found to be larger than P. m. maculosa from the Western Cape, having greater body mass, longer wings and a longer tail. Also, P. m. exultans of the eastern Free State and Lesotho was longer-winged and heavier than P. hypoxantha from north-eastern Free State, Mpumalanga and KwaZulu-Natal. Malachite Sunbirds Nectarinia formosa from populations at higher altitudes (Free State and KwaZulu-Natal) were larger (longer wings and heavier) than those occurring at lower altitudes in the Western Cape (De Swardt et al. 2004). Ashton (2002) reviewed body size data of several species and found that 76 of a total of 100 species showed a positive relationship between body size and latitude, and further concluded that species that occur at higher altitudes are larger than those in warmer areas. We also found that P. hypoxantha from the colder climate of the Drakensberg escarpment is larger than P. m. maculosa. This is consistent with Bergmann’s rule, which predicts that larger species are expected to be found in cooler regions (Ashton 2002; Blackburn, Gaston & Loder 1999).


Biometric data for both prinia species are represented by small samples in Maclean (1993), and only 20 males and 16 females of P. m. maculosa (possibly from the Western Cape) were recorded by Dean (2005a). The biometric data recorded during this study, including the measurement of the shorter rectrices (T5), and mass, represents the most detailed data set ever published for both P. maculosa and P. hypoxantha. Hockey, Dean & Ryan (2005) were the first to use biometric data and distribution maps of southern African birds on subspecific level, providing a useful reference guide for bird ringers. Chittenden, Allan & Weiersbye (2012) produced, for the first time, a field guide of southern African birds showing images of the different subspecies and distribution maps of selected species. The current work represents the first dataset for all three subspecies of P. maculosa (Dean [2005a] published data for P. m. maculosa only). This data will be a worthwhile addition to future updates of “Roberts birds of southern Africa”, to be used by ornithologists and bird ringers.
Kopij (2005) analysed the stomach contents of 20 P. maculosa and eight P. hypoxantha collected in the 1980s. During the present study a larger dietary sample for both prinia species was analysed. Prinias forage for insects on the stems and leaves of shrubs/bushes and between tufts of low-growing grass (Dean 2005a,b). Dean (2005a,b) and Kopij (2005) recorded coleopterans in prinia stomachs, consisting mainly of weevils (Curculionidae; both P. maculosa and P. hypoxantha), chrysomelid beetles (Chrysomelidae; P. maculosa) and monkey beetles (Scarabaeidae; P. hypoxantha), but ants (Hymenoptera) and flies (Hemiptera) were also important components of the diet of both species. Kopij (2005) also recorded a legless lizard (Acontidae) in the diet of the Karoo Prinia, and the Drakensberg Prinia was also observed feeding on the nectar of Aloe ferox (Dean 2005b). The reason for the low incidence of lepidopteran larvae in the diet, especially for P. hypoxantha, is probably due to soft items being more easily and rapidly digested, and therefore often being absent, in the digestive tract.
Our study revealed that small spiders (Araneae) are an important component in both species’ diets. Kopij (2005) did not record spiders in his study, although spiders and pseudoscorpions were recorded in the diet of the related Black-chested Prinia P. flavicans by Dean (2005c). Our data represents the first report of spider-eating by both P. maculosa and P. hypoxantha. The variety of insect orders represented in the diets of different prinia species suggests that they are opportunistic feeders, preying on most insects they encounter. It is therefore not surprising to report previously unrecorded arthropod taxa (e.g. Solifugae, Mantodea, Phthraptera, Orthoptera) in the diet of both species.
Kopij (2005) determined that various insectivorous passerines such as shrikes, bush-shrikes, tits and the smaller eremomela and crombec species included seeds in their diets. Because prinias forage for food by gleaning insects from vegetation (Dean 2005a,b), they probably also ‘accidentally’ ingest some seeds, but as many as 36.3% of P. hypoxantha had seeds in their stomachs, suggesting that for this species seed eating may be deliberate.
ACKNOWLEDGEMENTS
The authors are grateful to: Prof. Raurie Bowie (University of California, Berkeley, USA) for project ideas; Ruan Booysen for assistance with identification, sorting and weighing of stomach contents; Athol Marchant and Ian Rushworth (Ezemvelo KZN Parks) for permitting the capture and ringing of both species in Royal Natal National Park, Bergville and the Sani Pass area of Ukhalamba-Drakensberg Park; Johan & Sandra Taljaard (South African National Parks) for permission to capture and ring Karoo Prinias in Golden Gate Highlands National Park; Free State Department of Economic Development, Tourism and Environmental Affairs for permits to collect birds; various land owners in the Free State, Eastern Cape and Mpumalanga provinces who allowed access to their properties for field work; and several colleagues who assisted in ringing prinias at various localities: Arnold van der Westhuizen (Aliwal North and Rouxville areas), Morne Pretorius (Memel area), Jaco Smith (Royal Natal National Park), and ringers during Wakkerstroom ringing workshops for SAFRING and BirdLife South Africa. We thank Dr Mark Brown (Nature’s Valley Trust, Plettenberg Bay), Dr Penn Lloyd (Biodiversity Assessment & Management Pty Ltd, Australia) and Dr Robert Thompson (Percy FitzPatrick Institute of African Ornithology, University of Cape Town) for their thorough reviews.
REFERENCES
ASHTON, K.G. 2002. Patterns of within-species body size variation of birds: strong evidence for Bergmann’s rule. Global Ecology & Biogeography 11: 505–523.
BLACKBURN, T.M., GASTON, K. J. & LODER, N. 1999. Geographic gradients in body size: A clarification of Bergmann’s rule. Diversity and Distributions 5: 165–174.
CHITTENDEN, H, ALLAN, D.G. & WEIERSBYE, I. 2012. Roberts geographic variation of southern African birds. The John Voelcker Bird Book Fund, Cape Town, 284 pp.
CLANCEY, P. A. 1989. Four additional species of Southern African birds. Durban Museum Novitates 14: 140–152.
CLANCEY, P. A. 1997. Variation in the Karoo Prinia Prinia maculosa (Aves: Sylviidae) of South Africa. Durban Museum Novitates 22: 63–65.
DEAN, W.R.J. 2005a. Karoo Prinia Prinia maculosa. Pp. 845–846. In: HOCKEY, P.A.R., DEAN, W.R.J. & RYAN, P.G. (Eds). Roberts Birds of Southern Africa. Seventh edition. The Trustees of the John Voelcker Bird Book Fund, Cape Town.
DEAN, W.R.J. 2005b. Drakensberg Prinia Prinia hypoxantha. P. 846. In: HOCKEY, P.A.R., DEAN, W.R.J. & RYAN, P.G. (Eds). Roberts Birds of Southern Africa. Seventh edition. The Trustees of the John Voelcker Bird Book Fund, Cape Town.
DEAN, W.R.J. 2005c. Black-chested Prinia Prinia flavicans. Pp. 843. In: HOCKEY, P.A.R., DEAN, W.R.J. & RYAN, P.G. (Eds). Roberts Birds of Southern Africa. Seventh edition. The Trustees of the John Voelcker Bird Book Fund, Cape Town.
DE SWARDT, D.H., SYMES, C.T., DOWNS, C.T. & McFARLANE, M. 2004. An analysis of biometric data of the Malachite Sunbird Nectarinia famosa from five regions in South Africa. Afring News 33: 10–14.
DE SWARDT, D.H. 2010. Karoo and Drakensberg Prinias – the difference is in the spots! Culna 65: 16–17.
HOCKEY, P.A.R., DEAN, W.R.J. & RYAN, P.G. (Eds). 2005. Roberts Birds of Southern Africa. Seventh edition. The Trustees of the John Voelcker Bird Book Fund, Cape Town, 1296 pp.
KOPIJ, G. 2005. Diet of some insectivorous passerines in semi-arid regions of South Africa. Ostrich 76: 85–90.
MACLEAN, G.L. 1993. Roberts’ Birds of Southern Africa. John Voelcker Bird Book Fund, Cape Town, 871 pp.
RYAN, P.G. 2006. Family Cisticolidae (Cisticolas and allies). Pp. 378–490. In: DEL HOYO, J., ELLIOTT, A. & CHRISTIE, D.A. (Eds). Handbook of the birds of the world (vol. 11). Lynx Edicions, Barcelona.
SCHOLTZ, C.H. & E. HOLM. 1985. Insects of Southern Africa. Butterworths, Durban, 502 pp.
URBAN, E.K., FRY, C.H. & KEITH, S. (Eds). 1997. The birds of Africa, Volume 5, Thrushes to Puffback Flycatchers. Academic Press, London, 669 pp.



Comments are closed.