
Abstract
Forty-two reptile specimens from Mozambique are preserved in the collection of the National Museum, Bloemfontein, South Africa. This collection is comprised of 39 lizards referable to 10 families, 12 genera and 18 (possibly 19) species; and three snakes referable to two families, three genera and three species. Specimens were collected at 19 localities in six provinces (Tete, Manica, Sofala, Gaza, Inhambane and Maputo) in the central and southern parts of the country, mostly south of the Zambezi River. Morphological data on colour pattern, size and scalation are provided. Noteworthy new distribution records include the northern-most localities for Acontias aurantiacus aurantiacus (Peters) and Trachylepis depressa (Peters), the southern-most locality for Trachylepis megalura (Peters), and the second record of Cordylus rhodesianus (Hewitt) in Mozambique. A juvenile A. a. aurantiacus was found in the stomach of a large adult Trachylepis varia (Peters), representing the first record of an acontine lizard in the diet of any species of skink.
Key words: Reptiles, lizards, snakes, Mozambique, National Museum
INTRODUCTION
The reptile fauna of Mozambique consists of at least 294 species (Branch, Rödel & Marais 2005a,b; Schneider et al. 2005; Branch & Bayliss 2009; Jacobsen, Pietersen & Pietersen 2010; Bates & Broadley 2012; Portik et al. 2013a,b; Bates 2014; Branch, Bayliss & Tolley 2014; Reissig 2014; Conradie et al. 2016; Branch et al. 2017). Although numerous records are available for some parts of the country, many areas, especially in the north, remain poorly surveyed.
Records are available in several publications, including regional reptile assessments for southern Africa (lizards: FitzSimons 1943; snakes: FitzSimons 1962, Broadley 1983, 1990a), various taxonomic revisions (e.g. Broadley 1968, 1977a,b, 1980, 1998, 2000, 2001, 2002; Broadley & Gans 1978; Broadley & Broadley 1997; Broadley & Bauer 1998; Broadley & Branch 2002), and more recent papers for areas both south of the Zambezi River (e.g. Bates & Maguire 2009; Jacobsen, Pietersen & Pietersen 2010; Bates & Broadley 2012; Pietersen, Pietersen & Haacke 2013; Bates 2014; Pietersen 2014; Branch et al. 2017; Pietersen, Scholtz & Bastos 2018) as well as the more poorly known areas to the north thereof (e.g. Branch, Rödel & Marais 2005a,b; Branch & Bayliss 2009; Portik et al. 2013a,b; Branch, Bayliss & Tolley 2014; Conradie et al. 2016).
This paper provides details on all reptile specimens from Mozambique accessioned into the collection of the National Museum in Bloemfontein, South Africa. The taxonomic status of material is discussed; and details on size, colour pattern and scalation are provided. New records are compared to those in the published literature to determine their significance.
MATERIALS AND METHODS
Reptiles from Mozambique, preserved in 70% ethanol at the National Museum, were collected in 1989 (G.V. Haagner), 2007 (D. Maguire, S. Nielsen), 2008 & 2010 (J. Marais) and 2010 & 2011 (D. Pietersen). Lizards and snakes are the only kinds of Mozambican reptiles represented in the collection (also no amphibians).
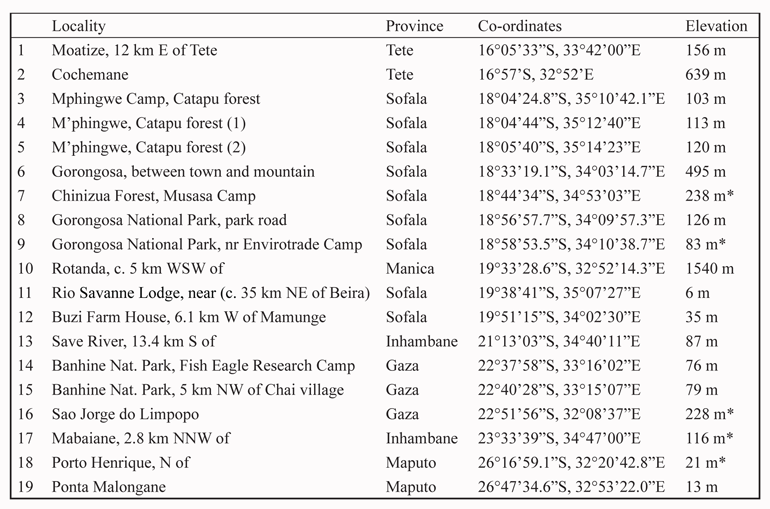
All localities are listed in Table 1, together with altitudes (meters above sea level) supplied by collectors (marked with an asterisk) or determined using Google Earth Pro (April–August 2018). The distribution of localities is indicated in Fig. 1.
Material was identified using descriptions and keys in the following publications: FitzSimons (1943); Broadley (1968, 1977a, 1980, 1990a, 2000, 2001, 2002); Broadley & Gans (1978); Broadley & Howell (1991), Jacobsen (1992); Broadley & Broadley (1997); Broadley & Branch (2002); Tilbury (2018) and Weinell & Bauer (2018).
Specimens were examined using a Nikon SMZ 745T stereo-microscope. Scale counts on the head and body are given for one side (if different on either side, this is reported). Measurements were performed using vernier callipers (0.01 mm) or a steel or plastic ruler (mm). Sex of snakes was determined by dissecting the right side of the tail base for presence (males) or absence (females) of an inverted hemipenis. Morphological character states important for identification are presented under ‘Morphology’ in the species accounts.
Families and species in the accounts below are ordered alphabetically within each suborder. The scincomorph families defined by Hedges (2014) are followed here. Common names, for the most part, follow Broadley (2001), Spawls et al. (2002) and Bates et al. (2014).
Table 1. Localities in Mozambique for reptiles in the National Museum (Bloemfontein), with provinces, coordinates, and elevation above sea level (m) as provided by the collectors (indicated by an asterisk) or determined using Google Earth Pro. Locality numbers correspond with numbers on the map (Fig. 1).
SAURIA
ACONTIDAE
Acontias aurantiacus aurantiacus (Peters, 1854)
Golden Legless Skink
Material. – Banhine National Park, 5 km NW of Chai village (NMB R8843: 118.1 mm snout to vent length [SVL] + 22.7 mm tail length = 140.8 mm total length); Banhine National Park, Fish Eagle Research Camp (NMB R8840: 128.2 + 25.2 = 153.4 mm; NMB R8841: 98.0 + 19.8 = 117.8 mm; NMB R8842: 123.5 + 25.1 = 148.6 mm); Save River, 13.4 km S of (NMB R10300: 158 + 25 = 183 mm; NMB R11686: juvenile from stomach of Trachylepis varia [Peters], NMB R10293).
Morphology. – Two azygous head shields posterior to rostral; scale rows at midbody 12; scale rows posterior to head shields similar to rest of body scales; frontal pentagonal, much larger than prefrontal; rostral usually bordered by seven shields, but six in NMB R11686 and eight in NMB R8842; prefrontal separated from loreals by frontonasals; supraoculars 2, supraciliaries 2, preocular 1, ocular 1, postocular 1 (in NMB R8842 the position and arrangement of these latter head shields appears to be aberrant). The Save River specimens are basically melanistic, but in the juvenile there is a cream stripe down the middle of the belly from throat to cloacal area, and in both specimens the chin is cream; the others are of the striped form, cream [in preservative] with a distinct pair of dark stripes over the middle of the back from nape to tip of tail, and a short stripe behind the eye extending onto the neck. Two specimens (NMB R8840–1) also have a feeble lateral stripe on either side extending to the tip of the tail, with an additional very feeble stripe on either side of the body posteriorly in NMB R8841; the dorsal part of the tail of the striped specimens has six dark stripes, those to the sides becoming progressively less distinct, and NMB R8841 also has a few vague stripes subcaudally.
Comments. – NMB R11686 is a juvenile that was found, in two parts, in the stomach of a Trachylepis varia (NMB R10293, see below).
This subspecies occurs in Mozambique south of the Save River and north of the Maputo and Pongolo Rivers, and in south-eastern Zimbabwe along the Nanetsi River (Broadley 1968, 1990b; Pietersen, Scholtz & Bastos 2018, Table 1). The new ‘13.4 km S of Save River’ locality is the northern-most record for this subspecies, the nearest other record being 65 km to the south-east at Inhassoro (Pietersen, Scholtz & Bastos 2018, Table 1).
Lamb, Biswas & Bauer (2010) showed that Typhlosaurus aurantiacus should be transferred to the genus Acontias Cuvier, and Pietersen, Scholtz & Bastos (2018) showed that two of the five subspecies (namely A. a. fitzsimonsi [Broadley 1968] and A. a. parietalis [Broadley, 1990b]) should be elevated to species status. Pietersen, Pietersen & Haacke (2013) noted that specimens of Typhlosaurus (= Acontias) from Banhine National Park may represent an undescribed species.
According to Broadley (1990b), A. a. aurantiacus is separated geographically from A. parietalis (considered a subspecies of T. aurantiacus by Broadley 1990b) in the south by the flood plains of the Maputo and Pongolo Rivers. Broadley (1990b) included only two Mozambican localities for A. parietalis, namely Inhaca Island and Ponta de Ouro, but his earlier specimens (as Typhlosaurus aurantiacus aurantiacus; Broadley 1968) from Lourenco Marques (= Maputo)—in the vicinity of Inhaca Island—are probably also referable to A. parietalis, and if so, then the southern-most record for A. a. aurantiacus is Chimonzo (see Broadley 1968).

AGAMIDAE
Agama armata Peters, 1854
Northern Ground Agama
Material. – Gorongosa National Park, park road (NMB R10280, female [no preanal pores]: 62.4 + 85.1 = 147.5 mm).
Morphology. – Dorsal scalation heterogeneous, with 10 more-or-less regular longitudinal rows (including two pairs of paravertebral rows) of enlarged spinose scales; dorsolateral scales with keels directed obliquely inwards to the vertebral line; gular with reticulate pattern of dark markings; ventral scales on the body and posterior part of the throat are weakly keeled; four large whitish, dark-edged, oval markings medially on the back between fore- and hindlimbs.
Comments. – Jacobsen (1992) distinguished Agama armata from Agama aculeata Merrem on the basis of its reticulate, rather than striped, gular pattern. He referred ground agamas from northern and central Mozambique to A. armata. The new record is situated about 70 km south-east of the nearest other Mozambique record (1833DA) plotted by Jacobsen (1992).
Agama mossambica Peters, 1854
Mozambique Agama
Material. – Chinizua Forest, Musasa Camp (NMB R10283, male [everted hemipenes]: 96.5 + 171.2 = 267.7 mm); Gorongosa, between town and mountain (NMB R10296, male [everted hemipenes]: 93.1 + 163.5 = 256.6 mm); Gorongosa National Park, near Envirotrade Camp (NMB R10277, female [no preanal pores]: 76.9 + 140.4 = 217.3 mm).
Morphology. – Dorsal scales more-or-less uniform, no distinctly enlarged scales; dorsolateral scales with keels directed obliquely inwards to the vertebral line; low vertebral crest on back; scales on back (especially paravertebrally) much larger than those on the belly; ventral scales on body and posterior part of throat are keeled (weakly on belly in NMB R10296); preanal pores 14 (NMB R10283) and 15 (NMB R10296).
Note: The key in FitzSimons (1943: 125) incorrectly treats this species as having “Dorsolateral scales on back with keels directed obliquely outwards [rather than inwards] from the vertebral line”.
Comments. – Found in lowland savannah and forest fringe in south-eastern Tanzania, Malawi, Mozambique as far south as the Buzi River, and eastern Zimbabwe (Broadley 1966; Branch 1998; Spawls et al. 2002; Portik et al. 2013; Conradie et al. 2016).
AMPHISBAENIDAE
Chirindia swynnertoni Boulenger, 1907
Swynnerton’s Worm Lizard
Material. – Buzi Farmhouse, 6.1 km W of Mamunge (NMB R10372: posterior part of body missing; NMB R10373: 110 mm SVL, tail broken/missing; NMB R10374: body twisted; NMB R10375: 131 + 14 = 145 mm).
Morphology. – Snout rounded; supralabials 2; segments in a midbody annulus 24 (midpoint estimated in NMB R10372 which had posterior part of body and tail missing); parietals not in lateral contact with post-supralabials; mental scale large, narrowing sharply behind, in contact with enlarged submedial scale (which is followed behind by 3–4 segments of the ‘second postgenial row’) in NMB R10374 & 10375, but separated by contact of first infralabials in NMB R10372 & 10373 (enlarged submedial scale followed behind by 3 segments); in NMB R10373 the enlarged submedial scale is flanked on either side by a smaller and narrower scale; no discrete scale (= postmental) between mental and enlarged submedial scale.
Comments. – This species occurs in the forests of Manicaland Province in eastern Zimbabwe and the mostly lowland provinces of Manica and Sofala in adjacent central Mozambique between latitudes 18°S and 21°S (Broadley & Gans 1978). The nearest locality to the new record is Amamba, 22 km SSW thereof (Broadley & Gans 1978). Measey & Tolley’s (2013) molecular analysis of sub-Saharan amphisbaenians indicated that the south-eastern African genus Chirindia Boulenger (samples of C. swynnertoni from Buzi were used) is most closely related to western African Cynisca Gray.
Zygaspis arenicola Broadley & Broadley, 1997 comb. nov.
Sand-dwelling Dwarf Worm Lizard
Material. – 2.8 km NNW of Mabaiane (NMB R10356: 71 + 13 = 84 mm, total length [TL]/tail length [tl] = 15.5%); Porto Henrique, N of (NMB R8703: 155 + 29 = 184 mm, TL/tl = 15.8%; NMB R8704: no measurements possible); Sao Jorge do Limpopo (NMB R9593: 155.6 + 32.7 = 188.3 mm, TL/tl = 17.4%).
Morphology. – Preoculars fused with prefrontal; postoculars 1; temporals 1; post-supralabials usually 0 (i.e. temporal and post-supralabial fused [= ‘temporal’], but one post-supralabial on either side in NMB R8703); body annuli + caudal annuli: 187 + 46 (NMB R10356), 198 + ? (NMB R8704), 201 + 48 (NMB R9593), 202 + 42 (NMB R8703); segments in a midbody annulus about 26 (NMB R10356 – difficult to count), 28 (NMB R8703–4), 30 (NMB R9593); pre-cloacal pores 4.
Comments. – The Porto Henrique specimens, found in humid soil, were reported on by Bates & Maguire (2009) and a tissue sample from NMB R8703 was used in the molecular analysis of Measey & Tolley (2013), while Pietersen (2014) discussed the other specimens. The Mabaiane specimen was collected in open mixed Julbernardia woodland. This locality fills a big gap between known localities of Z. v. arenicola to the north (nearest is Mapinhane, Mozambique, about 145 km to the NNE) and to the south (nearest is Chimonzo, Mozambique, 215 km to the SW) (Broadley & Broadley 1997). NMB R9593 from Sao Jorge do Limpopo was found in the top layer of sand and litter under a Wing Pod Tree (Xeroderris stuhlmanni [Taub.]) in open sandveld; this record is located 110 km SE of Broadley & Broadley’s (1997) nearest locality of Gonarezhou National Park in Zimbabwe (as noted by Pietersen 2014).
Like Acontias aurantiacus (see above), this species’ range in Mozambique appears to be restricted to the sands of the Mozambique Plain south of the Save River (Broadley & Broadley 1997). The Mabaiane locality is only 70 km north-west of Inhambane, the type locality of Zygaspis violacea (Peters) as restricted by Broadley & Broadley (1997). The latter species is known from only seven specimens from four mostly well separated coastal localities in southern Mozambique. There is some overlap in the scale counts and relative tail lengths used by the latter authors to distinguish between Z. v. arenicola and Z. violacea, suggesting that the status of these taxa should be investigated using a molecular analysis.
Broadley & Broadley (1997) noted that Z. violacea is the most derived species in the genus, exhibiting several features (fusion of various head shields) regarded as plesiomorphic. Zygaspis v. arenicola is almost identical in this regard, as most specimens similarly exhibit fusion of preoculars with prefrontals, parietals with frontals, a single postocular, and a single temporal (fused with post-supralabial); but Z. violacea also lacks a discrete parietal (Broadley & Broadley 1997). According to the latter authors, the latter two taxa differ in terms of scale counts as follows: body annuli 188–211 in Z. v. arenicola, 174–182 in Z. violacea; tail annuli 39–50 versus 50–59; segments in a midbody annulus 28–33 versus 34–40; tail length/total length 17.0–17.8% versus 18.9–23.3%. The Mabaiane specimen (NMB R10356) is similar to Z. violacea in terms of its head scalation and it even lacks parietals (see Fig. 13 in Broadley & Broadley 1997), but its body annuli count of 187, tail annuli count of 46, low number of segments to a midbody annulus (about 26), and short tail (15.5% of total length) place it with Z. v. arenicola. Broadley & Broadley (1997) noted that one of the syntypes (ZMB 50221) of Z. violacea from Inhambane also had 187 body annuli, and referred the specimen to Z. v. arenicola.
This opportunity was taken to consider the taxonomic status of Z. v. arenicola. The two subspecies of Z. vandami occur in different habitats: Z. v. vandami FitzSimons usually under stones or logs on sandy or humus-rich soils along the eastern escarpment of South Africa, while Z. v. arenicola is found in alluvial sands on the Mozambique Plain, extending into north-eastern KwaZulu-Natal, South Africa, north-eastern Swaziland, and inland to the Cretaceous sands of south-eastern Zimbabwe (Jacobsen 1989, as Z. violacea; Broadley & Broadley 1997; Litschka, Koen & Monadjem 2008). Typically, Zygaspis v. vandami has discrete anterior and posterior temporals, and anterior and posterior post-supralabials, as well as two postoculars, whereas Z. v. arenicola typically has the temporals and post-supralabials all fused into a single large shield, with a single elongated postocular (Broadley & Broadley 1997). While there is some variation to this head shield arrangement, most specimens of Z. v. arenicola exhibit considerable scale reduction (Broadley & Broadley 1997). Considering this morphological distinction and the geographical and habitat separation mentioned above, and in the context of a modern evolutionary species concept (Frost & Hillis 1990; De Quieroz 1998, 2007), I hereby formally propose that Zygaspis vandami arenicola Broadley & Broadley, 1997 be raised to full species status as Zygaspis arenicola Broadley & Broadley, 1997 comb. nov. It should be noted that in his checklist of the world’s amphisbaenians, Gans (2005: 7) elevated several known subspecies—including Z. v. arenicola—to species status, but this was done without justification and merely because “If the pattern and assignment of subspecific categories change again, it will be facile to derive and apply the pertinent information from this checklist.”
Zygaspis quadrifrons (Peters, 1862)
Kalahari Dwarf Worm Lizard
Material. – M’phingwe, Catapu forest (1) (NMB R9594: 170 + 36 = 206 mm); M’phingwe, Catapu forest (2) (NMB R9595: 177 + 36 = 213 mm).
Morphology. – Pair of discrete preoculars present, separating prefrontal from oculars; third supralabial bordered by a temporal and a post-supralabial; uniform brown above, paler (cream) below; body annuli + caudal annuli: 213 + 46 (NMB R9595) and 215 + 52 (NMB R9594).
Comments. – NMB R9594 was found under a log in a sand forest; NMB R9595 was in a small termitarium in dry deciduous forest. This species was recorded from the same general area (1835AA & AC) by Broadley & Broadley (1997).
CHAMAELEONIDAE
Chamaeleo dilepis Leach, 1819
Common Flap-neck Chameleon
Material. – Ponta Malongane (NMB R9481, male: 78 mm SVL).
Morphology. – Single series of enlarged white granules forms a gular crest on median line of throat, which continues along the belly to the vent as a (white) ventral crest; body scalation homogeneous, no enlarged tubercles; parietal crest distinct, not continuous with dorsal crest; casque slightly elevated posteriorly; tarsal process on foot (= male); occipital lobes small (about equal in length to distance that separates them from posterior commisure of mouth), non-rigid and separated; broad dark-edged pale stripe on each flank from behind forelimb insertion to a little beyond the middle of the body.
Comments. – This species is extremely widespread in sub-Saharan Africa, including much of Mozambique (Broadley 1966; Tilbury 2018), and has been recorded at Ponta do Ouro (Weinell, Portik & Bauer 2017) near the new locality of Ponta Malongane. Tilbury (2018) discussed variation in the characters defining the various putative subspecies of C. dilepis and suggested that, until a detailed molecular and morphological study of the complex has been conducted, it is best to consider this a single species with “geographically defined variants” (p. 377). The new specimen is clearly referable to the southern African (including southern half of Mozambique) form Chamaeleo dilepis quilensis Bocage, which is characterised by its small, non-rigid and separated occipital lobes. A molecular analysis by Main, Jansen van Vuuren & Tolley (2018) indicated that ‘quilensis’ may consist of three or more cryptic species, including a clade in KwaZulu-Natal (which may include neighbouring populations in Mozambique).
CORDYLIDAE
Cordylus rhodesianus (Hewitt, 1933)
Zimbabwean Girdled Lizard
Material. – Rotanda, c. 5 km WSW of [Chimanimani Mountains] (NMB R11073, male: 83.0 + 70.9 = 153.9 mm; NMB R11074, male: 79.5 + 73.5 = 153.0 mm).
Morphology. – Discrete loreal present; dorsal head shields ruffled, moderately rugose, no pronounced ridges bordering head shields; prefrontals narrowly separated by frontonasal in NMB R11074 but in contact posteriorly in NMB R11073; supralabials 6 (NMB R11074) or 7 (NMB R11073); infralabials 6 (left side infralabials fragmented in NMB R11073); interparietal large and separating anterior parietals in NMB R11074, but moderate and not separating them in NMB R11073; gular scales smooth (a few lateral scales with weak keels); lateral scales about as large as other dorsals, subcycloid with granular interspaces, keels on scales directed mostly obliquely upwards; nostril pierced in lower centre of nasal scale in NMB R11074, but nasal scales aberrant in NMB R11073; head depressed and expanded in temporal region; dorsal scales moderately keeled, ribbed, and often (NMB R11074) or occasionally (NMB R11073) finely serrated posteriorly; dorsals in 27 (NMB R11073) and 28 (NMB R11074) transverse rows, and 20 (NMB R11074) and 22 (NMB R11073) longitudinal rows; ventrals in 14 longitudinal rows; femoral pores per thigh 6 (also differentiated glandular femoral scales present = male); dorsum dark brown with large white spots and blotches, but NMB R11073 mostly with several narrow white crossbands on either side of the spine; underparts white, unmarked.
Comments. – The two specimens were found between rocks. This species is restricted to the eastern highlands of Zimbabwe, from Nyanga District southwards to the Chimanimani Mountains, where it extends into Mozambique. However, Broadley & Branch (2002) recorded only a single Mozambican specimen, from ‘Chimanimani Mountains’. The new material therefore represents only the second record of this species in Mozambique, and the first with detailed locality data.
EUGONGYLIDAE
Panaspis wahlbergi (A. Smith, 1849)
Wahlberg’s Snake-eyed Skink
Material. – Rio Savanne Lodge, near (NMB R10286: 35.8 + 55.0 = 90.8 mm).
Morphology. – Nostril pierced between two nasal scales, well separated from the frontal; interparietal distinct; frontoparietals fused; supraoculars 3; midbody scale rows 26; tail 1.5 times SVL. In life, top of head, back and tail grey-brown, flanks dark brown, underparts white with vivid orange at edges and on lower labials; narrow white dorsolateral and lateral stripes present (Fig. 2).
Comments. – A tissue sample from the specimen listed above (as ‘Beira’) was used in the molecular analysis of Medina et al. (2016). According to these authors, P. wahlbergi sensu stricto is restricted to the eastern half of South Africa and southern Mozambique (at least as far north as Rio Savanne area). Broadley’s (1966: 188) localities (as Ablepharus wahlbergi) of Beira and “8 miles NE of Beira” are near the new locality and probably also referable to this species/lineage (see Medina et al. 2016). The latter authors showed that Afroablepharus Greer is a junior synonym of Panaspis Cope.

Panaspis sp. (clade ‘Mozambique 1’, Medina et al. 2016)
Western Mozambique Snake-eyed Skink
Material. – Gorongosa National Park, near Envirotrade Camp (NMB R10288: 28.0 + 20.8r = 48.8+ mm).
Comments. – A tissue sample from the juvenile specimen listed above, as ‘SVN 693’ from ‘Gorongosa National Park’, was used in the analysis of Medina et al. (2016). The specimen is part of a lineage that probably represents an undescribed species. It is part of a consignment of loaned material and is thus not currently available for re-examination. Broadley’s (1966) record (as Ablepharus wahlbergi) of Gorongosa Mountain is probably referable to this lineage.
GEKKONIDAE
Hemidactylus mabouia (Moreau de Jonnes, 1818)
Common Tropical House Gecko
Material. – Moatize, 12 km E of Tete (NMB R6507, female: 51 + 66 = 117 mm; NMB R6508, female: 45 + 45 = 90 mm).
Morphology. – Dorsum with small, carinate (usually 2–3 keels), subimbricate to juxtaposed granules, including about 12 longitudinal rows of enlarged conical tubercles; original tail depressed; distance from end of snout to anterior border of eye greater than distance from posterior border of eye to posterior border of ear opening; about nine (NMB R6507) or 10 (NMB R6508) transverse dorsal scale rows in a proximal caudal verticel; subdigital lamellae on fourth toe in six pairs with a single terminal pad; no preano-femoral pores (females).
Comments. – Currently considered to have a cosmopolitan range (Branch 1998), and widespread in Mozambique (Broadley 1977a). However, ongoing genetic analyses indicate that cryptic radiation may have occurred in the H. mabouia species complex in south-eastern Africa, including Mozambique, and this may require taxonomic re-interpretation (A. Bauer pers. comm., June 2018).
Hemidactylus platycephalus Peters, 1854
Flat-headed Tree Gecko
Material. – Gorongosa National Park, near Envirotrade Camp (NMB R10278, female: 75.7 + 87.4 = 163.1 mm).
Morphology. – Dorsum with smooth, subimbricate to juxtaposed granules, including about 10 longitudinal rows of moderate sized and weakly conical tubercles; original tail depressed; distance from end of snout to anterior border of eye greater than distance from posterior border of eye to posterior border of ear opening; about 14 transverse dorsal scale rows in a proximal caudal verticel; subdigital lamellae on fourth toe in seven pairs with a single terminal pad; no preano-femoral pores (female).
Comments. – This is a mainly East African species, with a widespread distribution in Mozambique (almost as far south as Maputo) where it is commonly sympatric with H. mabouia (Broadley 1977a). It has been recorded from Bunga Inselberg in the north-western part of Gorongosa National Park (Branch et al. 2017).
Lygodactylus capensis capensis (A. Smith, 1849)
Common Dwarf Gecko
Material. – Gorongosa National Park, near Envirotrade Camp (NMB R10279, male: 28.0 + 35.6 = 63.6 mm); Mphingwe Camp, Catapu forest (NMB R10281, male: 30.7 mm SVL, tail tip missing).
Morphology. – Inner digit of hands and feet minute, with tiny claw; mental with a pair of posterior clefts; subcaudal scales subequal, no rows of enlarged scales; preanal pores 5 (NMB R10279) and 7 (NMB R10281); pale, dark-edged, dorsolateral stripe on either side of body from behind eye, but weakly developed and on anterior part of body only in NMB R10281.
Comments. – This widespread subspecies was recorded from Gorongosa Mountain by Broadley (1966), and Branch et al. (2017) recorded ‘Lygodactylus cf. capensis’ from Bunga Inselberg in the north-western part of Gorongosa National Park. Lygodactylus c. capensis has been recorded in sympatry with Lygodactylus chobiensis FitzSimons at Catapu Concession (Pietersen 2014).
GERRHOSAURIDAE
Gerrhosaurus flavigularis Wiegmann, 1828
Yellow-throated Plated Lizard
Material. – Chinizua Forest, Musasa Camp (NMB R10282: 121 + 310 = 431 mm).
Morphology. – Scales on soles of feet smooth; supraoculars 4; supraciliaries 5; dorsals in 61 transverse rows (from post-nuchal row to row above vent) and 22 longitudinal rows; ventrals in 8 longitudinal rows; SVL/head length [tip of snout to upper part of ear opening on right side of head] = 5.4; distinct white, black edged, dorsolateral stripe on either side of body.
Comments. – This widespread species appears to be absent from a large part of the Mozambique Plain (Broadley 1966; Branch 1998). The molecular analysis of Bates et al. (2013) indicated some sub-structuring within G. flavigularis; the taxonomic status of a lineage of east African populations (including samples from Bazaruto and the Zambezi Delta in Mozambique) requires further investigation.
LYGOSOMIDAE
Mochlus sundevallii (A. Smith, 1849)
Sundevall’s Writhing Skink
Material. – Gorongosa National Park, park road (NMB R10299: 63.1 mm SVL, tail broken/missing).
Morphology. – Prefrontals distinct; midbody scale rows 28; dorsal surface of body and tail brown with numerous white spots and extensive white barring on the sides.
Comments. – The specimen was found inside a log that was broken open. A molecular and morphological analysis by Freitas et al. (2018) indicated that Mochlus afer (Peters) is a junior synonym of M. sundevallii. The former was distinguished from the latter by its larger size and a black-and-white speckled dorsum (see Broadley & Howell 1991), but Freitas et al. found that although some specimens could be distinguished in this way, such differences may be due to heterochronic morphological shifts rather than signifying species boundaries. Records for M. sundevallii and ‘M. afer’ indicate that this species is widespread in Mozambique (Broadley 1966; Branch 1998; Portik et al. 2013; Conradie et al. 2016).
MABUYIDAE
Trachylepis depressa (Peters, 1854)
Eastern Sand Skink
Material. – Chinizua Forest, Musasa Camp (NMB R10290: 70.9 + 96.1 = 167. 0 mm).
Morphology. – Plantar and subdigital scales non-spinose, smooth and tuburcular; dorsal scales with
seven keels each; midbody scale rows 31; a single row of (non-granular) scales above window in eyelid; anterior border of ear opening with three lobules, two lanceolate, the lowermost one triangular; head and body strongly depressed; top of back plain brown, distinct black stripe from behind eyes to hindlimbs, with an indistinct pale lateral stripe below it; underparts cream.
Comments. – In Mozambique this species is found only south of the Zambezi River, but its range extends into south-eastern Zimbabwe and the north-eastern parts of South Africa (Broadley 1966; FitzSimons 1943; Broadley 2000; Bates et al. 2014). Broadley (1966) recorded it from only nine localities in Mozambique (Beira, Caia, Dondo, Inhaca Island, Masieni, Portuguese Island, Rikatla, Santa Carolina Island, Savane) and noted that the type locality of ‘Tette’ is probably incorrect as these skinks are restricted to the alluvium of the Mozambique Plain. Broadley (1990b, 1992) latter recorded this species from various Mozambican islands in the Bazaruto Archipelago as well as Inhaca Island, while Jacobsen, Pietersen & Pietersen (2010) recorded it from the San Sebastian Peninsula. Most known localities are coastal and there do not appear to be many records for Sofala and Manica Provinces (see map in Branch 1998).
Broadley (1966) referred Cott’s (1934) Mabuia lacertiformis (Peters) record of ‘Caia’ to T. depressa, but treated it as somewhat dubious as the specimen/s were not preserved and thus not available for examination. Cott (1934) did not provide a description of the material, and although Caia seems too far east for Trachylepis lacertiformis (Peters) and too far north for Trachylepis punctulata (Bocage) (see Broadley 1975), it is within range of other skinks such as Trachylepis varia (see Broadley 1966; Branch 1998; Weinell & Bauer 2018) with which it could have been confused. The new Chinizua Forest record is 110 km south-west of Caia, and 100 km north-north-east of Dondo, the nearest other record to the south. It represents the most northerly confirmed record (specimens available for examination) of this species.
Trachylepis megalura (Peters, 1878)
Grass-top Skink
Material. – Rio Savanne Lodge, near (NMB R10291: 53.4 mm SVL, tail broken/missing).
Morphology. – Plantar and subdigital scales non-spinose, smooth and tubucular; dorsal scales smooth; midbody scale rows 26; ear opening distinctly elongated (antero-posteriorly) and without lobules. In life, the top of the head, back and at least the base of the tail are grey (back mostly brown in preservative), the flanks are brown with a narrow white stripe from under the eye extending to the hindlimbs, and the venter is white (Fig. 3).
Comments. – This species ranges from Ethiopia southwards to eastern and central Africa, but appears to be patchily distributed in some areas (Broadley 2000; Spawls et al. 2002). Broadley (1986: 30) recorded it from “Chiniziua District (1835Cd)” and noted that this locality represented a south-westerly range extension of 725 km from Lumbo (the only Mozambican locality listed by Broadley 1966) on the north coast of Mozambique. The Chiniziua specimen was the first of its species found south of the Zambezi River. The Rio Savanne specimen reported on here is only the second known specimen from south of the Zambezi, and the southern-most record for the species. It represents a range extension of about 90 km south-south-west of the Chiniziua record.

Trachylepis striata (Peters, 1844)
Striped Skink
Material. – Chinizua Forest, Musasa Camp (NMB R10284: 86.3 + 141.1 = 227.4 mm); Cochemane (NMB R6512: 51 mm SVL, tail broken/missing; mass 4.1 g).
Morphology. – NMB R10284: Plantar scales spinose; subdigital lamellae with median keel and lateral carinations; subocular scale excluded from lip; dorsal scales on fourth toe in two rows on the proximal phalanx; skin not fragile; head and body not strongly depressed; dorsal scales with three distinct keels each; midbody scale rows 34; lamellae under fourth toe 21 (left foot), 22 (right); broad dorsolateral stripe (2–2.5 scales wide at midbody) on either side of body from above eyes to base of tail; underparts white with grey speckles on throat; prefrontals in contact; anterior border of ear opening with four small, roundish lobules. NMB R6512: Not available for re-examination.
Comments. – This species is widespread in Mozambique, including several recorded localities in the vicinity of Chinizua Forest (Broadley 1966, 1977b, 2000; Portik et al. 2013; Conradie et al. 2016).
Trachylepis varia (Peters, 1867)
Eastern Variable Skink
Material. – Gorongosa National Park, near Envirotrade Camp (NMB R10289: 55.0 + 57.1r = 112.1+ mm); Rio Savanne Lodge, near (NMB R10292: 46.6 + 81.6 = 128.2 mm); Save River, 13.4 km S of (NMB R10293, female [ovaries examined]: 63.3 + 94.0 = 157.3 mm).
Morphology. – Plantar scales spinose; subdigital lamellae tricarinate; subocular narrowed below (i.e. with a slight anterior prolongation above); dorsal scales on fourth toe in a single row almost to the base; dorsal scales with three distinct keels, but in NMB R10293 several scales have an additional short weak keel on either side; midbody scale rows 31 (NMB R10289, 10293) and 32 (NMB R10292); anterior border of ear opening with 2–3 short, bluntly pointed, subtriangular lobules; nostril pierced laterally in a nasal scale and directed outwards and upwards, its centre behind the rostro-labial suture (only slightly so in NMB R10293), separated from first supralabial by a distance much less than its own diameter; lower border of subocular over half length of upper; posterior loreal distinctly longer than anterior loreal; distinct white lateral stripe on either side of body, from behind nasal scale to hindlimb; distinct narrow, pale, dorsolateral stripe on either side of body in NMB R10289 and 10293, but in NMB R10292 the pale dorsolateral stripes are vague and there is also a pale, vague median (vertebral) stripe on the back; dorsal surface of back with scattered dark blotches (also scattered white specks in NMB R10289), but few in NMB R10293; flanks with scattered small pale spots.
Comments. – NMB R10293, a large female, contained a juvenile Acontias a. aurantiacus (NMB R11686, in good condition generally but in two pieces) in its stomach which it appears to have swallowed tail-first (Fig. 4). The anterior part of the stomach contained the larger portion of the prey, with its head positioned near the junction of oesophagus and stomach. It seems unlikely that the T. varia bit the Acontias into two pieces, so the possibility exists that it was found in that condition before being consumed. Trachylepis varia preys mainly on insects and other invertebrates, but occasionally other lizards are eaten (FitzSimons 1943; Broadley 1966; Jacobsen 1982; Branch 1998; Spawls et al. 2002). The lizards referred to above appear to be in reference to Broadley (1966: 167) who noted that a T. varia from Mutare in Zimbabwe regurgitated an adult Panaspis wahlbergi “nearly as long as itself”; and Jacobsen (1982: 99) who found tails of this same prey species (P. wahlbergi) in stomachs of two adult T. varia from Nylsvley Nature Reserve in Limpopo Province, South Africa, and also referred to an adult T. varia (3.1 g) from the same locality that regurgitated the “head and body” of an adult “Panaspis”. It should be noted here that according to the map in Weinell & Bauer (2018), the Nylsvley population, and possibly even the Mutare one, may be referable to Trachylepis damarana (Peters). This new dietary record apparently represents the first record of an acontine lizard in the diet of any species of skink.
The molecular analysis of Weinell & Bauer (2018) indicated that T. varia sensu stricto occurs throughout Mozambique, Swaziland and eastern South Africa, while the range of the central-southern African species Trachylepis damarana extends into western Mozambique. Broadley (1966) listed several localities for T. varia in Mozambique.

SERPENTES
COLUBRIDAE
Thelotornis mossambicanus Bocage, 1895
Eastern Twig Snake
Material. – Buzi Farmhouse, 6.1 km W of Mamunge (NMB R10361, female: 345 + 187+ [tail tip missing] = 532+ mm).
Morphology. – Specimen appears largely faded, but top of head and temporal region largely uniform (minor traces of darker markings); indications of dark markings on neck; supralabials 8 (4th & 5th enter orbit); infralabials 11; postoculars 3; ventrals 152; paired subcaudals 132+ (tail tip missing); anal shield divided; midbody scale rows 19.
Comments. – This is the only species of Thelotornis A. Smith recorded from this area by Broadley (2001).
LAMPROPHIIDAE
Prosymna stuhlmannii (Pfeffer, 1893)
East African Shovel-snout
Material. – Chinizua Forest, Musasa Camp (NMB R10285, female: 91.2 + 16.2 = 107.4 mm).
Morphology. – Dorsal scales mostly pale with dark edges; tip of head (especially rostral and internasal) mostly cream and noticeably paler that rest of body; scales on back smooth; nostril pierced in a semi-divided nasal; pupil subelliptic; loreal present; preoculars 1; postoculars 2; internasal a broad band; ventrals 134; paired subcaudals 32; anal shield undivided; midbody scale rows 15.
Comments. – This species is widely distributed in Mozambique. The nearest known locality is ‘Muda-Lamego’ (Broadley 1980), probably a point about halfway between the two villages by those names, i.e. about 87 km south-west of the Chinizua Forest locality.
Psammophis orientalis Broadley, 1977
Eastern Striped-belly Sand Snake
Material. – Gorongosa National Park, near Envirotrade Camp (NMB R10287, female: 663 + 324+ [tip of tail missing] = 987+ mm).
Morphology. – Back mostly dark but scales with pale centers, broad cream dorsolateral stripe on either side extending over most of the body and tail; venter with a pair of distinct black stripes from the neck to the end of the tail where it fades out, pale yellow between stripes and cream on either side; supralabials 8 (4th and 5th enter orbit); infralabials 9 (first four on either side in contact with pair of anterior sublinguals); preoculars 1; postoculars 2; one elongate loreal; post-nasal entire and without posterior prolongation; ventrals 155; paired subcaudals 91+ (tip of tail missing); anal shield undivided; midbody scales rows 17.
Comments. – Psammophis subtaeniatus orientalis was elevated to species status by Broadley (2002). It occurs in East Africa from southern Kenya to Tanzania, Malawi, eastern Zimbabwe and as far south as Pomene in southern Mozambique; recorded from several localities in the vicinity of Gorongosa National Park (Broadley 2002).
DISCUSSION
This paper documents a collection of 42 reptile specimens from Mozambique, preserved in the collection of the National Museum, Bloemfontein, South Africa. The collection is comprised of 39 lizards referable to seven families, 12 genera and 18 species (excluding an undescribed, candidate species of Panaspis); and three snakes referable to two families, three genera and three species. Specimens were collected at 19 localities in six provinces (Tete, Manica, Sofala, Gaza, Inhambane and Maputo) in the central and southern parts of the country, mostly south of the Zambezi River.
Particularly noteworthy records are the northern-most localities for the lizards Acontias a. aurantiacus and Trachylepis depressa, and the southern-most locality for Trachylepis megalura. The record ‘13.4 km S of Save River’ represents a 65 km north-westerly extension of the range of A. a. aurantiacus; the ‘Musasa Camp, Chinizua Forest’ locality is a 100 km NNE extension of the range of T. depressa; while the ‘near Rio Savanne Lodge’ record for T. megalura extends this species’ known distribution by about 90 km SSW of the nearest other record. Two specimens of Cordylus rhodesianus represent only the second record of this lizard species in Mozambique, and the first with detailed locality data. A record of the snake Prosymna stuhlmannii from the Chinizua locality mentioned above fills a large gap in this species’ known range in Mozambique.
The stomach of a large adult Trachylepis varia collected near the Save River contained a juvenile A. a. aurantiacus. This represents the first record of an acontine lizard in the diet of any species of skink.
It is hoped that the information above will make this collection and its associated data accessible and useful to other researchers, and contribute towards the eventual compilation of a detailed checklist of the reptiles of Mozambique.
ACKNOWLEDGEMENTS
I thank Luke Verburgt (Enviro-Insight CC) for preparing the distribution map; and the two referees, Mr Stephen Spawls (Norwich, England) and Dr Shelley Edwards (Rhodes University, Grahamstown), for their valuable comments on the manuscript.
REFERENCES
BATES, M.F. 2014. Geographical distributions: Tetradactylus africanus. African Herp News 61: 35–36.
BATES, M.F. & BROADLEY, D.G. 2012. Geographical distributions: Cordylus vittifer. African Herp News 56: 34–35.
BATES, M.F. & MAGUIRE, D. 2009. Geographical distribution: Zygaspis vandami arenicola. African Herp News 47: 43–44.
BATES, M.F., TOLLEY, K.A., EDWARDS, S., DAVIDS, Z., DA SILVA, J.M. & BRANCH, W.R. 2013. A molecular phylogeny of the African plated lizards, genus Gerrhosaurus Wiegmann, 1828 (Squamata: Gerrhosauridae), with the description of two new genera. Zootaxa 3750 (5): 465–493.
BATES, M.F., BRANCH, W.R., BAUER, A.M., BURGER, M., MARAIS, J., ALEXANDER, G.J. & DE VILLIERS, M.S. (Eds). 2014. Atlas and Red List of the Reptiles of South Africa, Lesotho and Swaziland. Suricata 1. South African National Biodiversity Institute, Pretoria, pp. i–xvii + 1–485.
BRANCH, W.R. 1998. Field Guide to Snakes and other Reptiles of Southern Africa. Third Edition. Struik Publishers, Cape Town, pp. 399.
BRANCH, W.R. & BAYLISS, J. 2009. A new species of Atheris (Serpentes: Viperidae) from northern Mozambique. Zootaxa 2113: 41–54.
BRANCH, W.R., BAYLISS, J. & TOLLEY, K. 2014. Pygmy chameleons of the Rhampholeon platyceps complex (Sauria: Chamaeleonidae): Description of four new species from isolated ‘sky islands’ of nor-thern Mozambique. Zootaxa 3814 (1): 1–36.
BRANCH, W.R., GUYTON, J.A., SCHMITZ, A., BAREJ, M.F., NASKRECKI, P., FAROOQ, H., VERBURGT, L. & RÖDEL, M.-O. 2017. Description of a new flat gecko (Squamata: Gekkonidae: Afroedura) from Mount Gorongosa, Mozambique. Zootaxa 4324 (1): 142–160.
BRANCH, W.R., RÖDEL, M.-O. & MARAIS, J. 2005a. Herpetological survey of the Niassa Game Reserve, northern Mozambique – Part I. Reptiles. Salamandra 41 (4): 195–214.
BRANCH, W.R., RÖDEL, M.-O. & MARAIS, J. 2005b. A new species of rupicolous Cordylus Laurenti 1768 (Sauria: Cordylidae) from Northern Mozambique. African Journal of Herpetology 54 (2): 131–138.
BROADLEY, D.G. 1966. The Herpetology of South-East Africa. Ph.D. dissertation, University of Natal, Pietermaritzburg, pp. 680.
BROADLEY, D.G. 1968. A revision of the African genus Typhlosaurus Wiegmann (Sauria: Scincidae) Arnoldia Rhodesia 3 (36): 1–20.
BROADLEY, D.G. 1977a. A review of the Hemidactylus mabouia complex in southeastern Africa (Sauria: Gekkonidae). Arnoldia Rhodesia 8 (19): 1–15.
BROADLEY, D.G. 1977b. A review of the Mabuya striata complex in South-East Africa (Sauria: Scincidae). Occasional Papers of the National Museums and Monuments of Rhodesia B6 (2): 45–79.
BROADLEY, D.G. 1980. A Revision of the African snake genus Prosymna Gray (Colubridae). Occasional Papers of the National Museums and Monuments of Rhodesia B6 (7): 481–556.
BROADLEY, D.G. 1983. FitzSimons’ Snakes of Southern Africa. Delta Books, Johannesburg, 376 pp.
BROADLEY, D.G. 1986. Geographical distribution: Mabuya megalura. Journal of the Herpetological Association of Africa 32: 30.
BROADLEY, D.G. 1990a. FitzSimons’ Snakes of Southern Africa. Revised edition with addendum. Delta Books, Johannesburg, 376 pp. + Addendum.
BROADLEY, D.G. 1990b. The herpetofaunas of the islands off the coast of south Mocambique. Arnoldia Zimbabwe 9 (35): 469–493.
BROADLEY, D.G. 1992. Reptiles and amphibians from the Bazaruto Archipelago, Mozambique. Arnoldia Zimbabwe 9 (38): 539–548.
BROADLEY, D.G. 1998. A review of the African Elapsoidea semiannulata complex (Serpentes: Elapidae). African Journal of Herpetology 47 (1): 13–23.
BROADLEY, D.G. 2000. A review of the genus Mabuya in southeastern Africa (Sauria: Scincidae). African Journal of Herpetology 49 (2): 87–110.
BROADLEY, D.G. 2001. A review of the genus Thelotornis A. Smith in eastern Africa, with the description of a new species from the Usambara Mountains (Serpentes: Colubridae: Dispholidini). African Journal of Herpetology 50 (2): 53–70.
BROADLEY, D.G. 2002. A review of the species of Psammophis Boie found south of Latitude 12° S (Serpentes: Psammophiinae). African Journal of Herpetology 51 (2): 83–119.
BROADLEY, D.G. & BAUER, A.M. 1998. A review of the Mabuya quinquetaeniata complex in East Africa (Sauria: Scincidae). African Journal of Herpetology 47 (2): 43–58.
BROADLEY, D.G. & BRANCH, W.R. 2002. A review of the small east African Cordylus (Sauria: Cordylidae), with the description of a new species. African Journal of Herpetology 51 (1): 9–34.
BROADLEY, D.G. & BROADLEY, S. 1997. A revision of the African genus Zygaspis Cope (Reptilia: Amphisbaenia). Syntarsus 4: 1–24.
BROADLEY, D.G. & GANS, C. 1978. Southern forms of Chirindia (Amphisbaenia, Reptilia). Annals of Carnegie Museum 47 (3): 29–51.
BROADLEY, D.G. & HOWELL, K.M. 1991. A check list of the reptiles of Tanzania, with synoptic keys. Syntarsus 1: 1–70.
CONRADIE, W., BITTENCOURT-SILVA, G.B., ENGELBRECHT, H.M., LOADER, S.P., MENEGON, M., NANVONAMUQUITXO, C., SCOTT, M. & TOLLEY, K.A. 2016. Exploration into the hidden world of Mozambique’s sky island forests: new discoveries of reptiles and amphibians. Zoosystematics & Evolution 92 (2): 163–180.
COTT, H.B. 1934. The Zoological Society’s Expedition to the Zambezi, 1927: No. 5. On a Collection of Lizards, mainly from Portuguese East Africa, with Descriptions of new Species of Zonurus, Monopeltis, and Chirindia. Proceedings of the Zoological Society of London 1: 145–173, plates I–III.
DE QUIEROZ, K. 1998. The General Lineage Concept of Species, Species Criteria, and the Process of Speciation. A Conceptual Unification and Terminological Recommendations. Pp. 55–75. In: Howard, D.J. & Berlocher, S.H. (eds), Endless forms: species and speciation. Oxford University Press, U.S.A.
DE QUIEROZ, K. 2007. Species Concepts and Species Delimitation. Systematic Biology 56 (6): 879–886.
FITZSIMONS, V. 1943. The lizards of South Africa. Memoirs of the Transvaal Museum (Pretoria) 1: 1–528.
FITZSIMONS, V.F. 1962. Snakes of Southern Africa. Purnell & Sons, Cape Town, 423 pp.
FREITAS, E.S., BAUER, A.M., SILER, C.D., BROADLEY, D.G. & JACKMAN, T.R. 2018. Phylogenetic and morphological investigation of the Mochlus afer-sundevallii species complex (Squamata: Scincidae) across the arid corridor of sub-Saharan Africa. Molecular Phylogenetics and Evolution 127: 280–287.
FROST, D.R. & HILLIS, D.M. 1990. Species in concept and practice: Herpetological applications. Herpetologica 46 (1): 87–104.
GANS, C. 2005. Checklist and bibliography of the Amphisbaenia of the world. Bulletin of the American Museum of Natural History 289: 1–130.
HEDGES, S.B. 2014. The high-level classification of skinks (Reptilia, Squamata, Scincomorpha). Zootaxa 3765 (4): 317–338.
JACOBSEN, N.H.G. 1982. The ecology of the reptiles and amphibians in the Burkea africana ‒ Eragrostis pallens savanna of the Nylsvley Nature Reserve. M.Sc. dissertation, University of Pretoria, Pretoria, pp. xvii + 257.
JACOBSEN, N.H.G. 1989. A herpetological survey of the Transvaal. Unpubl. Ph.D. dissertation. University of Natal, Durban.
JACOBSEN, N.H.G. 1992. The status of Agama aculeata armata Peters 1854, (Reptilia: Agamidae). Journal of the Herpetological Association of Africa 41: 30–34.
JACOBSEN, N.H.G., PIETERSEN, E.W. & PIETERSEN, D.W. 2010. A preliminary herpetological survey of the Vilanculos Coastal Wildlife Sanctuary on the San Sebastian Peninsula, Vilankulo, Mozambique. Herpetology Notes 3: 181–193.
LAMB, T., BISWAS, S & BAUER, A.M. 2010. A phylogenetic reassessment of African fossorial skinks in the subfamily Acontinae (Squamata: Scincidae): evidence for parallelism and polyphyly. Zootaxa 2657: 33–46.
LITSCHKA, T., KOEN, C. & MONADJEM, A. 2008. Geographical distribution: Zygaspis vandami arenicola. African Herp News 46: 24–25.
MAIN, D.C., JANSEN VAN VUUREN, B. & TOLLEY, K.A. 2018. Cryptic diversity in the common flap-necked chameleon Chamaeleo dilepis in South Africa. African Zoology 53 (1): 11–16.
MEASEY, G.J. & TOLLEY, K. 2013. A molecular phylogeny for sub-Saharan amphisbaenians. African Journal of Herpetology 62 (2): 100–108.
MEDINA, M.F., BAUER, A.M., BRANCH, W.R., SCHMITZ, A., CONRADIE, W., NAGY, Z.T., HIBBITS, T.J., ERNST, R., PORTIK, D.M., NIELSEN, S.V., COLSTON, T.J., KUSAMBA, C., BEHANGANA, M., RÖDEL, M-O. & GREENBAUM, E. 2016. Molecular phylogeny of Panaspis and Afroablepharus skinks (Squamata: Scincidae) in the savannas of sub-Saharan Africa. Molecular Phylogenetics and Evolution 100 (2016): 409–423.
PIETERSEN, D.W. 2014. New distribution records of herpetofauna in Mozambique south of the Zambezi River, including additional records from Parque Nacional de Banhine. Annals of the Ditsong National Museum of Natural History 4: 174–180.
PIETERSEN, D.W., PIETERSEN, E.W. & HAACKE, W.D. 2013. First herpetological appraisal of the Parque Nacional de Banhine, Gaza Province, southern Mozambique. Annals of the Ditsong National Museum of Natural History 3: 153–163.
PIETERSEN, D.W., SCHOLTZ, C.H. & BASTOS, A.D.S. 2018. Multi-locus phylogeny of Southern African Acontias aurantiacus (Peters) subspecies (Scincidae: Acontinae) confirms the presence of three genetically, geographically and morphologically discrete taxa. Zootaxa 4442 (3): 427–440.
PORTIK, D.M., MULUNGU, E.A., SEQUEIRA, D. & McENTEE, J.P. 2013a. Herpetological Surveys of the Serra Jeci and Namuli Massifs, Mozambique, and an Annotated Checklist of the Southern Afromontane Archipelago. Herpetological Review 44 (3): 394–406.
PORTIK, D.M., TRAVERS, S.L., BAUER, A.M. & BRANCH, W.R. 2013b. A new species of Lygodactylus (Squamata: Gekkonidae) endemic to Mount Namuli, an isolated ‘sky island’ of northern Mozambique. Zootaxa 3710 (5): 415–435.
REISSIG, J. 2014. Girdled Lizards and their relatives: Natural History, Captive Care and Breeding. Edition Chimaira, Frankfurt am Main, 249 pp.
SCHNEIDER, M.F., BURAMUGE, V.A., ALIASSE, L. & SERFONTEIN, F. 2005. Checklist and Centres of Vertebrate Diversity in Mozambique. Forestry Department and Eduardo Mondlane University, Maputo.
SPAWLS, S., HOWELL, K., DREWES, R. & ASHE, J. 2002. A field guide to the reptiles of East Africa (Kenya, Tanzania, Uganda, Rwanda and Burundi). Academic Press, San Diego, 543 pp.
TILBURY, C.R. 2018. Chameleons of Africa: An Atlas including the Chameleons of Europe, the Middle East and Asia. Second Edition. Frankfurt Contributions to Natural History, Volume 77. Edition Chimaira, Frankfurt am Main, 643 pp.
WEINELL, J.L. & BAUER, A.M. 2018. Systematics and phylogeography of the widely distributed African skink Trachylepis varia species complex. Molecular Phylogenetics and Evolution 120 (2018): 103–117.
WEINELL, J.L., PORTIK, D.M. & BAUER, A.M. 2017. Noteworthy records of amphibians and reptiles from Mozambique. African Herp News 65: 3–10.



Comments are closed.